Conceiving of Body Awareness in Terms of Modality and Modularity
Abstract
Abstract
Studies in animal self-awareness is a most promisimg direction in the field of evolutionary and comparative psychology. Different experimental methods were developed and tested provoding a vast amount of contradictory data. These data gave rise to a lot of difficulties and discordance in theoretical justification of the phenomenon of self-representation. We propose a model of body representation in animals, within the framework of which the self-representation is viewed not as a sort of monolithic abstract knowledge of an individual about the self, but as a set of modality-specific subsystems. The purpose of such dedicated systems is to process various physical characteristics of the individual’s body as related to solving specific behavioral problems. Based on the provisions of this approach, a number of requirements are formulated for the methods aimed at studying self-awareness in animals. Also, using the ideas of the Differentiation-Integration approach, the main trends in the evolution of self-representation may be identified.
Представление о Телесном Осознании в Терминах Модальности и Модульности
Хватов И.А. а, Харитонов А.Н. б*
аНаучно-образовательный центр Биопсихологических исследований Московского института психоанализа, 121170 Москва, Россия ORCID iD: 0000-0002-6939-244X
бИнститут психологии Российской академии наук, 129366 Москва, Россия, ORCID iD: 0000-0002-4801-9937
Аннотация: За последнюю четверть века изучение Self-awareness стало одним из наиболее бурно-развивающихся научных направлений в области Animal cognition. Было разработано и апробировано множество различных экспериментальных методик, накоплен обширный объем противоречивых данных, что породило немало трудностей в теоретическом осмыслении феномена саморепрезентации у животных. Мы предлагаем оригинальную теорию репрезентации собственного тела животными, в рамках которой саморепрезентации представляется не в качестве единого абстрактного знания индивида о себе, но в виде множество модально специфических модулей, учитывающих различные физические характеристики тела индивида для решения частных поведенческих задач. На основе положений развиваемой теории формулируется ряд требований к методикам, нацеленным на изучение Self-awareness у животных. С привлечением дифференционно-интеграционного подхода выделяются основные тенденции эволюции саморепрезентации.
Ключевые слова: самопрезентация, представление о собственном теле, модальность самопрезентации, модули самопрезентации, самосознание, осознание тела, зеркальное самопознание, дифференционно-интеграционный подход.
Информация об авторах
Хватов Иван Александрович, кандидат психологических наук, заведующий кафедрой общей психологии, руководитель Научно-образовательного центра бмопсихологических исследований, НОЧУ ВО Московский институт психоанализа, г. Москва, Российская Федерация, ORCID: https://orcid.org/0000-0002-6939-244X, e-mail: ittkrot1@gmail.com
Харитонов Александр Николаевич, кандидат психологических наук, старший научный сотрудник, Инсттитут психологии РАН, г. Москва, Российская Федерация, ORCID: https://orcid.org/0000-0002-4801-9937, e-mail: ankhome47@list.ru
Introduction
Self-representation is a mental representation of an individual’s own qualities, which is necessary for regulating their behavior and interaction with the environment. This concept denotes an extremely wide range of diverse phenomena: self-concept (Rogers, 1994; Gore, Cross, 2011); self-image (Fein, Spencer, 1997); self-schema (Markus, 1977), self-esteem (Hewitt, 2009), social identity (Turner, Oakes, 1986), etc. In humans, most of these types and/or forms of self-representation are culturally and socially determined (Stolin, 1983).
Which form of self-representation is the phylo- and ontogenetically earliest? In answering this question, one may be guided by the fundamental principle: the initial mental images in terms of development contain a minimum of abstractions, being direct representations of specific objects, i.e. material bodies (Zorina, Poletaeva, 2003). Accordingly, the earliest (most probably, the starting point), both in phylo- and ontogenesis, should be considered the mental representation of one’s own body, an individual’s idea of the physical qualities of own body necessary to regulate his behavior and interaction with the environment (Khvatov et al., 2023). This phenomenon has been discussed for a long time. W. James called it the “physical self” (1911), and V.V. Stolin (1983) proposed the idea of “taking oneself into account”. Nevertheless, the theoretical development of such a seemingly simple concepts is still far from complete.
Currently, two forms of representation of one’s own body are distinguished, i.e. body schema and body image. Body schema is an unconscious (implicit), sensorimotor representation that arises directly in the course of behavior, which is the basis for constructing an egocentric coordinate system (Head & Holmes, 1911; Keizer et al., 2013). A significant part of the research in the body schema and the possibilities of its modification is carried out on people (Johnson-Frey, 2004). The body schema is highly differentiated and flexible, integrating into its structure external objects that are in physical contact with the individual, for example, the tools used by him (Carlson et al., 2010; Carlson, 2013; Costantini et al., 2014; Garbarinia et al., 2015; Gozli & Brown, 2011; Ritchie & Moeller et al., 2016). In accordance with the change in the body scheme, there is also a modification of the subjective perception of the surrounding reality, i.e. the egocentric coordinate system (Barabanschikov, 2002; Berti & Frassinetti, 2000; Giglia et al., 2015; Costantini et al., 2014).
Body image is a conscious (explicit) representation of one’s own body characteristics (e.g., one’s own appearance), which serves as the basis for constructing an allocentric coordinate system (Schilder, 1935). This phenomenon is often studied clinically. Distortions of body image are observed in various psychiatric and neurological disorders (Denes, 1999; Artoni et al., 2021).
Apart from psychology, distinction between body schema and body image has been the basis of much work in neuropsychology, cognitive neuroscience and philosophy (de Vignemont, 2010; Dijkerman & de Haan, 2007; Di Vita et al., 2016; Gallagher, 1986; Paillard, 1999). The body schema functions directly in the process of behavior, whereas body image concerns the abstract imagination of a complex of one’s own physical properties. This distinction is consistent with the general functional hypothesis according to which perception and action require different transformations of sensory signals, follow different rules and are thus serviced by different cortical and subcortical pathways. This has been shown for visual and auditory signaling (Dyson, Dunn, & Alain, 2010; Hall, 2003; Milner & Goodale, 1995; Milner & Goodale, 2008; Romanski et al., 1999; Warren, Wise, & Warren, 2005; Zatorre et al., 2002) as well as for somatosensory systems (Dijkerman, de Haan, 2007). However, in most everyday situations, the “perceived body” does not confront the “acting body”. In this regard, it has been argued that the discussed difference between body schema and body image is putative and these two phenomena act as different functional mannifestations of individual’s representation of one’s own body (Pitron & de Vignemont, 2017).
A completely different narrative emerged in the fields of evolutionary and comparative psychology that study the phylogeny and species-specific diversity of forms of body representation in animals. Firstly, in this case the authors do not focus on the differences between the body scheme and the body image. There are only isolated studies of the body scheme in animals in the field of neuroscience (Maravita, & Iriki, 2004). This is probably due to the fact that most experiments conducted on animals we discuss below are behavioral, that is, directly related to the body scheme, and therefore it is not possible to isolate the body image only. Secondly, to designate self-representation, as applied to animals, in the English-language literature the researchers mostly use the term “self-awareness”, that is the ability of an individual to perceive himself (primarily the body) separately from the characteristics of the external world and/or other individuals (Ferris, 2012).
We have deliberately avoided using the term “self-awareness” so far, since there are currently disagreements in cognitive science, which is dominant throughout the world. Among the proponents of the Cultural Activity Approach which is widely spread in the post-Soviet space there is also much controversy regarding its applicability to animals.
In the English-language literature, authors operate with the concept of “self-awareness” rather loosely identifying its features not only in humans, but also in a huge number of different species (de Waal, 2019; Gallup & Anderson, 2020; Hunt, 2004). It should be mentioned here that promotion of “animal consciousness” is also driven by the interests of bioethics (The Cambridge Declaration on Consciousness, 2012; The New York Declaration on Animal Consciousness, 2024), which has no direct relation to science. So we will allow ourselves to leave this aspect without comment.
Unlike in cognitive psychology, the Cultural Activity Approach has a clearer understanding of consciousness, considering it as a level of mental evolution specific to humans (Filippova, 2012). In this regard, Russian comparative psychology is not so much concentrated on animal consciousness, as on the phylogenetic prerequisites of human consciousness (Zorina & Poletaeva, 2003). This topic has been developed in detail in the works of Russian psychologists (Fabri, 2004; Filippova, 2012; Leontiev, 1981). A.N. Leontiev defines consciousness as a reflection of objective reality in which its objective stable properties are distinguished regardless of the subject’s attitude towards it. The impetus for the formation of consciousness is placed on labor as a specific collective activity, involving the division of functions between several participants, which, in turn, determines the emergence of language generated by human relationships with other people that develop in the process of labor (Leontiev, 1981). Consequently, self-awareness is considered one of the key components of consciousness as the highest level of phylo- and ontogenetic development of the mind inherent in humans and distinguishing them from other animal species (Filippova, 2012; Leontiev, 1981), even when it comes to such a relatively simple phenomenon as awareness of one’s own body.
In our opinion, the current terminological dispute is due to the differences in the theoretical foundations of the two approaches. Whereas cognitive psychology is predominantly based on positivism and neo-positivism, inherited from behaviorism, the Cultural Activity Approach is based on the sociologically oriented ideology of dialectical materialism. Without going into further details of methodological discussions, we should admit that the question of emergence of the representation of one’s own body (as the basis for other types and forms of self-representation) in the course of phylogenesis remains open and attracts much attention.
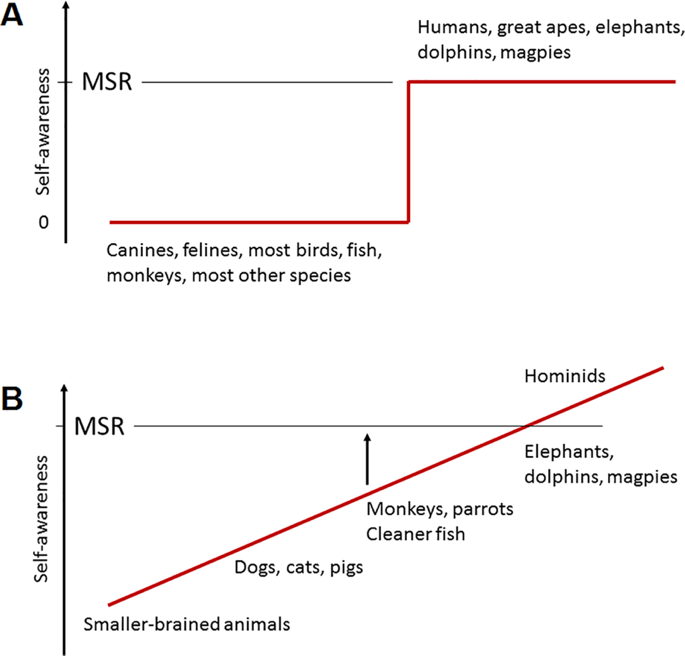
Figure 1. Two models of the evolution of self-representation (English: “self-awareness”): A – binary, B – gradualistic (see text for explanations) (from de Waal, 2019).
Currently there are two most widely adopted models that are relevant to explain the emergence of representation of one’s own body in animals in the process of mental evolution: binary and gradualistic (Fig. 1).
- The binary model postulates sudden emergence of self-representation, the presence of which can be judged by passing the mirror self-recognition (MSR) test. In 1970, Gordon Gallup Jr. experimentally investigated the possibility of self-recognition in a mirror in two naive males and two naive females of wild chimpanzees (Pan troglodytes). Each chimpanzee was first placed in an experimental room for two days. Then a full-length mirror was placed in the room for 80 hours. Various behavioral patterns were then recorded during the interaction of the monkeys with the mirror. Initially, the chimpanzees made threatening gestures towards their own reflections, supposedly seeing them as a threat in return. By the end of the experiment, the chimpanzees used their own reflections to guide self-directed behaviors, such as grooming parts of their body not previously observed without a mirror, nose picking, and the like. Gallup extended the study by manipulating the chimpanzees’ appearance and observing their reactions to the mirror reflection (Gallup, 1970). Gallup lightly anesthetized the chimpanzees and then applied an alcohol-soluble dye to the edge of the eyebrow and the upper half of the opposite ear. When the dye dried, it had virtually no olfactory or tactile properties. Gallup then returned the chimpanzees to the cage (with the mirror removed) and they regained full consciousness. He then recorded the frequency with which the chimpanzees spontaneously touched the marked areas of their skin. After 30 minutes, the mirror was reintroduced into the room and the frequency of touching the marked areas was again registered. The frequency of touching the marked areas of the body was four to ten times higher in the presence of a mirror than in its absence.
Based on the results obtained, it was concluded that the chimpanzees remembered their appearance and noticed changes in it, and also understood that the image in the mirror was equivalent to their own body.
To date, the ability to recognize oneself in a mirror has been established in humans (starting from the age of two years) (Beulah, 1972), great apes (Allen, 2007; Povinelli et al., 1993; Suarez & Gallup, 1981; Walraven et al., 1995), dolphins (Reiss & Marino, 2001), killer whales (Delfour & Marten, 2001), Asian elephants (Plotnik et al., 2006), Eurasian magpies (Prior et al., 2008), Indian house crows (Buniyaadi et al., 2020), cleaner wrasses (Kohda et al., 2019; 2023), and Atlantic crabs (Robinson, 2023). On the other hand, many animals failed the MSR test, i.e. sea lions (Hill et al., 2015), giant pandas (Ma et al., 2015), dogs (Coren, 2001), monkeys (Hauser et al., 2001; Ma et al., 2015; Rajala et al., 2010; Roma et al., 2007; Shaffer, & Renner, 2000), grey parrots (Hill et al., 2015), great tits (Kraft et al., 2017), cichlid fish (Hotta et al., 2018), and octopuses (Mather & Kuba, 2013).
- The gradualistic model assumes that all species have self-awareness, as it is necessary for survival. Rochart identifies 5 levels of self-representation development in human ontogenesis (Rochat, 2003). Later Frans de Waal extrapolated this concept to phylogenesis (de Waal, 2019). From this point of view, representation of one’s own body can be imagined as an onion, i.e. the phenomenon is multifaceted, it becomes more complex and supplemented as the mind evolves. However, the highest developmental level of self-awareness is associated with passing the MSR test.
Thus, the above models, in one way or another, both assume that the highest level of development of the representation of one’s own body is expressed in the ability to recognize oneself in a mirror or in a photo.
One should note that there is much criticism of the MSR approach. Firstly, it cannot be applied to animals whose visual modality is not the leading one (most mammals) (Coren, 2004). Secondly, both animals and humans may not show interest in marks on their own bodies so far as they may not recognize this mark as something abnormal (Fox, 1982; Asendorpf et al., 1996). Thirdly, some animals may feel uncomfortable looking at “the other” in the mirror, since this provokes aggression (Anderson, 1984; Couchman, 2011). Fourthly, vertical reflective surfaces are almost never found in nature (Lei, Yanyu, 2023).
A persistent idea widely spread in scientific circles is that the ability to recognize oneself in a mirror, firstly, is an expression of complex high-level cognitive processes, and secondly, indicates the inclusion of the subject into flexible social connections and predisposition to understand the intentions of conspecifics, or “theory of mind” (Krachun et al., 2019; Lei, Yanyu, 2023). Meanwhile, the ability to pass the appropriate test is demonstrated by those animals whose mental regulation of behavior is extremely simple (however not primitive), i.e. fish (Kohda et al., 2019; 2023) and crabs (Robinson, 2023).
In summary, it should be concluded that at present the field of evolutionary and comparative studies of representation of own body in animals is in crisis signaled by the impossibility of consistently explaining the genesis and evolution of this phenomenon, as well as its species diversity. This crisis is caused by a number of both theoretical and methodological contradictions.
Below we propose an alternative model of the genesis and evolution of the representation of one’s own body by animals, which, in our opinion, can provide a basis for a consistent integration of the diverse empirical data in this area, as well as propose an efficient method for studying the phenomenon under discussion.
Results
2. A model of modal-modular representation of one’s own body
The model of representation of one’s own body by animals that we propose is based on two provisions.
- In the process of mental regulation of behavior any individual must take into account the physical characteristics of the body (boundaries, volume, mass) and correlate them with the physical characteristics of external objects. In other words, animals need to “take themselves into account” (Stolin, 1983). This thesis is well substantiated by the concept of “embodied cognition” (Varela et al., 1991), which is being rapidly developed in modern cognitive psychology. The basic tenet of this concept is that the mind matures, exists and develops as a result of the interaction of the integral physical body (not just the brain or nervous system) with physical objects in the external environment, including social contacts. In particular, this means that cognition is rooted in the environment, it unfolds in the context of the surrounding world. Even autonomous (separated from the environment) cognition is based on the individual’s bodily experience (Wilson, 2002).
To develop an original model of self-representation based on the above provisions of Embodied Cognition Approach, we will need to clarify the most basic categories.
The mind here is considered as an information based process that regulates the behavioral interaction of an individual with the diversity of objects in the environment. In turn, behavior is a form of life activity of an individual (a unitary organism), providing adaptation by changing spatial relationship with objects of the environment by means of locomotor and manipulatory acts. In this context, a unitary organism as integral and indivisible entity (Marfenin, 2016) is opposed to a modular, decentralized one. Accordingly, many animals (but for sessile or parasitic), as well as heterotrophic protists, belong to unitary creatures: the mind is necessary for them to regulate their own behavior. It is reasonable to note that in the course of the discussion we inevitably come into contact with the problem of the genesis of the mind. However, a detailed analysis of this topic is not the purpose of this article, although it certainly deserves rethinking in the light of modern facts from the field of biology (Khvatov, 2011; Lyon, 2015; Lyon et al., 2021).
It is important to emphasize that the definition of the mind presented above is not radically new. Firstly, it is based on the traditional understanding of information as a process of removing uncertainty (Shannon, 1948). Secondly, this definition is entirely consistent with the classical Soviet psychology consideration of the mind through the prism of its involvement in the process of regulation of activity and behavior.
Accordingly, for the behavioral satisfaction of its own need, the animal each time creates subjective representations of a variety of external objects, as well as a representation of itself. Thus, the mind is located “on the edge” of two objective realities, external and internal, the reality of one’s own organism (Zinchenko, 1999). By constructing and synthesizing these representations, the individual builds an image of a holistic situation, the event (joint existence) (Barabanshchikov, 2002), thus providing a basis for behavioral planning. Kurt Lewin called this phenomenon the Psychological Field of Life Space (Lewin, 2000). For the possibility of behavioral satisfaction of an actual need, the synthetic structure of the image of the event must be harmonious in the sense of a complementary relationship between the representations of one’s own body and the surrounding objects. As a result, the planned set of locomotor and manipulatory acts is embodied in accordance with the physical limitations and affordances (Gibson, 1966; Norman, 2013) of both the individual’s body and the characteristics of current situation.
The problem arises in a situation when a behavioral act cannot be realized due to the presence of limitations of one’s own body and/or material environment. In such a case, the individual either discovers this circumstance at the planning stage, or makes a mistake, trying unsuccessfully to perform the behavioral act.
Then one of three possible scenarios is possible:
- an individual’s refusal to satisfy his needs behaviorally;
- an individual’s reorganization of the mental image of an event, taking into account the existing physical limitations of one’s own body and surrounding objects, in order to plan a new behavioral act;
- a systematic physical modification of one’s own body, surrounding objects, or both, in order to give them the required qualities for the possibility of implementing the original behavioral act.
The latter two options provide the basis for planning various psychological experiments to study the peculiarities of animal thinking (Zorina, Poletaeva, 2003; Fabri, 2004). As an example of the second option, we can cite the classic experiment to discover the detour (Fabri, 2004). To illustrate the third option, it is appropriate to mention Wolfgang Kohler’s experiments with chimpanzees. In this case, by constructing a tower of boxes, the chimpanzee modified the environment and, by extending his arm with a stick, he modified his own body.
It follows from the above that in any ethological experiment, in the process of problem solving when planning its behavior, an animal uses the representation of its own body in some way. The problem is that, for a researcher building a model of the animal’s mental processes on the basis of the obtained empirical data, it is often extremely difficult to isolate the self-representation, i.e. the representation of the parameters of animal’s own body, and separate it from the representation of the parameters of other surrounding objects. In other words, changing an independent variable we often cannot say for sure whether the tested animal takes into account its body or other objects when it changes its behavior. A better assumption would be it takes into account both, as its body and the external world in subjective representation do not exist in isolation from each other. This circumstance should be considered when designing experiments aimed at identifying the features of the representation of one’s own body by animals of certain species.
2. The second assumption concerns the structural organization of the representation of one’s own body. Here one may rely on the methodology of Grounded Cognition (Barsalou, 2008; 2010). Cognitive psychology traditionally represents cognition as a sort of computation occurring in modular systems that process abstract and amodal content. According to grounded cognition theory, cognition is based on modality-specific and bodily-conditioned processes. Modality is understood not as a sensory quality, but as the specificity of individual’s behavior within certain conditions when solving a specific problem. Accordingly, grounded cognition appears to be associated with the ideas of enactivism (Thompson, 2010), which proposes to consider a mental image not as a simple copy (representation) of objective reality, but as a system continuously constructed by an individual in the process of his behavioral activity, and therefore dependent on specificity of the latter. In turn, enactivism, which emerged in the 90ies of the past century, was in a way anticipated by the basic provisions of the Theory of Activity (Leontiev, 1975).
Bringing the above ideas into the framework of the model we are constructing suggests that the representation of one’s own body by an individual is not a monolithic system of storage of abstract information about all sorts of characteristics of oneself. We propose the self-representation is a set of subsystems modally specialized in relation to the individual’s mental regulation of his behavior under specific material conditions that require consideration of clearly defined parameters of his body.
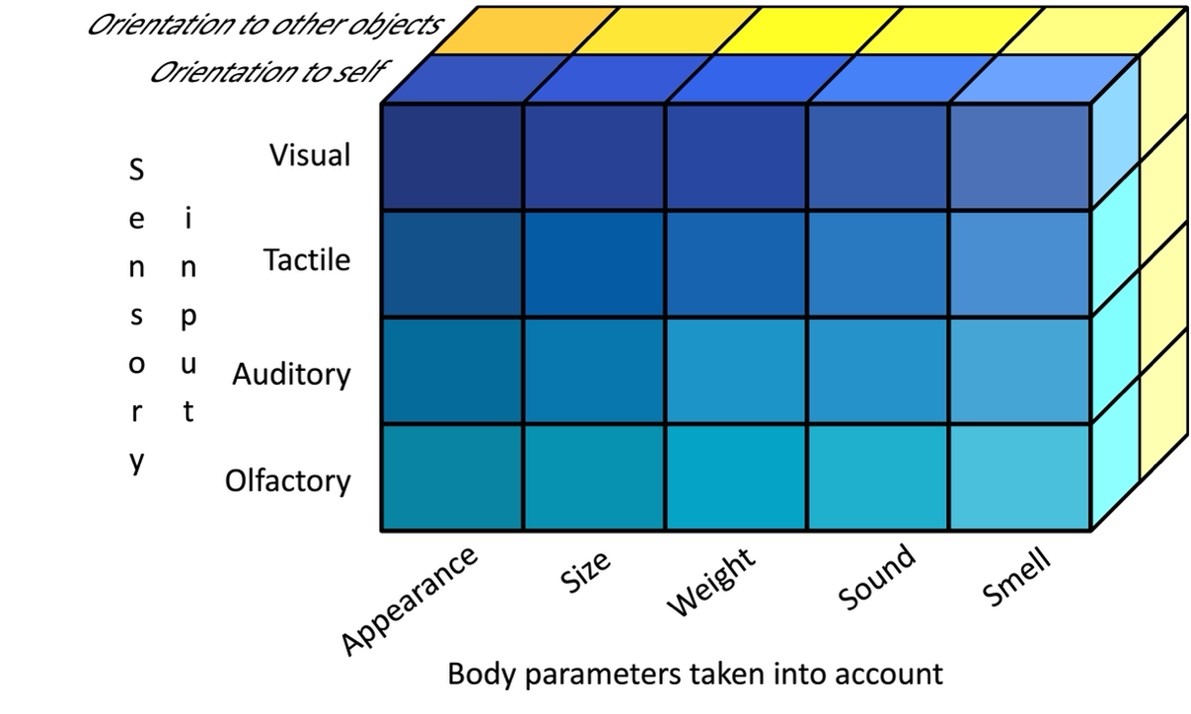
Figure 2. Three modalities of representation of one’s own body (see text for explanations)
To further discuss the model, we introduce and analyze two concepts: the modality of self-representation and the module of self-representation. The modality of self-representation is a particular dimension of the representation of one’s own body by an individual arising in accordance with the specific features of behavioral activity and the ecological context within which this representation is a simple copy (representation) of objective reality, but as a system continuously constructed by an individual in the process embodied. We distinguish three modalities: sensory input, the parameters of one’s own body taken into account, and the motivational orientation of behavior within which self-representation functions (Fig. 2).
Sensory input is a specific quality of sensory data used to construct self-representation, something that A.N. Leontiev called “sensory fabric” (Leontiev, 2024). Here we distinguish four types of sensory inputs: visual, tactile, auditory and olfactory (Fig. 2) engaged in self-representation. It is important to note that this typology serves only as a theoretical starting point so far as it is possible to distinguish several more variants of sensory inputs in different animal species (e.g., electroreception, magnetoreception, etc.).
By the parameters of one’s own body we mean specific manifestations of the individual’s body taken into account by an individual when constructing a self-representation in the context of current behavior. To begin with, it is necessary to distinguish five types of such parameters: one’s own appearance, one’s own body size, one’s own body weight, one’s own body sound (both voice and other sounds that occur during locomotor and/or manipulative acts), and one’s own body odor. For example, when choosing a hole of a suitable size for penetration, an animal must take into account the size (boundaries) of its body, and when determining the strength of the support for passage its weight. At the same time, one should not confuse sensory input with the parameters of one’s own body taken into account. Naturally, under certain conditions they may coincide. Alternatively, looking at one’s own reflection in the mirror, we construct a representation of one’s own appearance based on the visual modality. On the other hand, a person, determining the penetrability of a certain opening (say, a doorway) with his own body and, accordingly, taking into account the size of the latter, also relies on visual data (Warren & Whang, 1987). A rat, due to weak development of its vision, solving a similar problem, is primarily guided by tactile sensations from its vibrissae (Krupa et al., 2001).
The third modality, motivational orientation, is subdivided into only two types and is determined according to whether the individual’s behavior is oriented toward self-study and/or self-modification (e.g., cleaning one’s body) or not. We shall discuss this modality in more detail, since its types affect two main methodological approaches currently used to study self-representation in animals.
If the subject is focused on self-study and/or self-modification (Fig. 2, blue blocks), we assume that his or her own body is a goal. The ability to represent one’s body for the purpose of self-modification is studied using the MSR method described earlier. In Gallup’s experiment (Gallup, 1970), chimpanzees performed mark-directed behavior by touching bright marks on their faces that were visible only in the mirror. A similar approach, but in a different modality, is implemented in the so-called “olfactory mirror” method aimed at identifying the ability of animals to distinguish their odor traces from other odors, including conspecifics. To date, dogs (Horowitz, 2017) and gray wolves (Gatti et al., 2020), snakes (Burghardt, et al., 2021) and geckos (Szabo, Ringler, 2023) have passed the test. In particular, R. Cazzolla Gatti notes “scent-rolling behavior” in wolves (Gatti et al., 2020).
When the behavior is oriented to other objects (Fig. 2, yellow-orange blocks), the subject’s own body (and consideration of its parameters) is not the goal, but the means. This manifestation of self-representation is studied using the “Body-as-obstacle” task. The experimental design presupposes that in order to solve an external task (for example, getting bait), the animal needs to consider the parameters of its body (size, weight, etc.), which creates an obstacle for implementation of the goal-directed behavior. The ability to consider one’s own weight was revealed in children aged 18-26 months (Brownell et al. 2007) and Indian elephants (Dale, Plotnik, 2017), domestic dogs (Lenkei et al., 2021) and rats (Khvatov, Kharitonov, 2021). The ability to take into account the size of their body was discovered in children aged 18–26 months (Brownell et al. 2007), budgerigars (Schiffner et al., 2014), domestic dogs (Lenkei et al., 2020), snakes (Khvatov et al., 2019), gray crows (Khvatov et al., 2021), domestic ferrets (Khvatov et al., 2023), snakes (Khvatov et al., 2019), toads (Khvatov & Zhelankin, 2018) and rats (Khvatov et al., 2024).
We believe that the representation of one’s own body when orienting individual behavior toward other objects is a basic type in the above-mentioned modality. This means that both in the phylogenetic and ontogenetic development, animals first acquire the ability to identify the parameters of their own body considering them as possible obstacles to the implementation of their behavior, and only under this condition that they can have motivation for purposeful self-study
and/or self-modification. Accordingly, from the point of view of reveiling the features of self-awareness, the body-as-obstacle method has greater power in comparison with MSR. This thesis may be confirmed by the fact that in the overwhelming majority of cases of using the body-as-obstacle task, positive results were obtained: animals took into account the parameters of their own body, even if not on the first try, but were able to learn (e.g. toads, see: Khvatov & Zhelankin, 2018; and snakes (Khvatov et al., 2019)). In addition, the MSR task is more complex than direct consideration of one’s own body parameters, since it involves instrumental activity. The subject must indirectly interact with the body using the mirror as a tool, and this involves additional cognitive mechanisms.
The colored blocks in Fig. 3 represent the modalities of own body representation studied to date in animals of different species. In addition to the above data, the ability of humans to recognize their own voices (Kaplan et al., 2008) and mirror-induced self-directed behavior in mice (Yokose et al., 2023) were established.

Figure 3. Modalities of body representation studied to date in animals of different species (for explanations see the text).
The selection of modalities of self-representation of one’s own appearance based on the olfactory modality or the representation of one’s own smell based on the auditory modality. For a further grounding of the model, we additionally introduce the category of “self-representation module”. The self-representation module is a subsystem, i.e. a relatively autonomous structural unit of representation of one’s own body, specialized in solving a number of similar behavioral problems and formed by means of integration of several different modalities. Accordingly, the modules of self-representation correspond to the properties identified by J. Fodor (1983), in particular, domain specificity, informational encapsulation, and fast speed of data processing. On the other hand, such a definition of the self-representation module is close to the understanding of “a functional organ” by A.A. Ukhtomsky (Ukhtomsky, 1927) and “a perceptual system” within the framework of the ecological approach (Gibson, 1979).
In turn, the evolutionary emergence of various self-representation modules occurs as a result of the mechanism of exaptation (Gould & Vrba, 1982) of the functions of individual sensory and effector systems of animals in accordance with the changing features of their ecology and behavior. Accordingly, different animal species may have different (both qualitatively and quantitatively) sets of self-representation modules, similarly to a Swiss army knife that contains many various tools (a metaphor proposed by Cosmides and Tooby, 1994). Further we consider several examples of such subsystems.
One of the most obvious illustrations will be humans. We have a highly integrated (in terms of its modal structure) visual-tactile-auditory module of representation of our own body, providing great behavioral flexibility and a high degree of abstraction of self-awareness (Fig. 4). This thesis is substantiated by the following. First, a person has a visual-tactile synthesis: visual and tactile systems may provide similar cognitive processes to sensory inputs that enable humans to rely merely on one modality in the absence of another to recognize surrounding objects (Tabrik et al., 2021). Similarly, it has been demonstrated that visual recognition of appearance itself and recognition of one’s own voice have similar neural support (Kaplan et al., 2008). Secondly, a high correlation has been found between the results of the MSR and body-as-obstacle task tests in humans in the second half of the second year of life (Barth et al., 2004; Brownell et al., 2007; Moore et al., 2007).
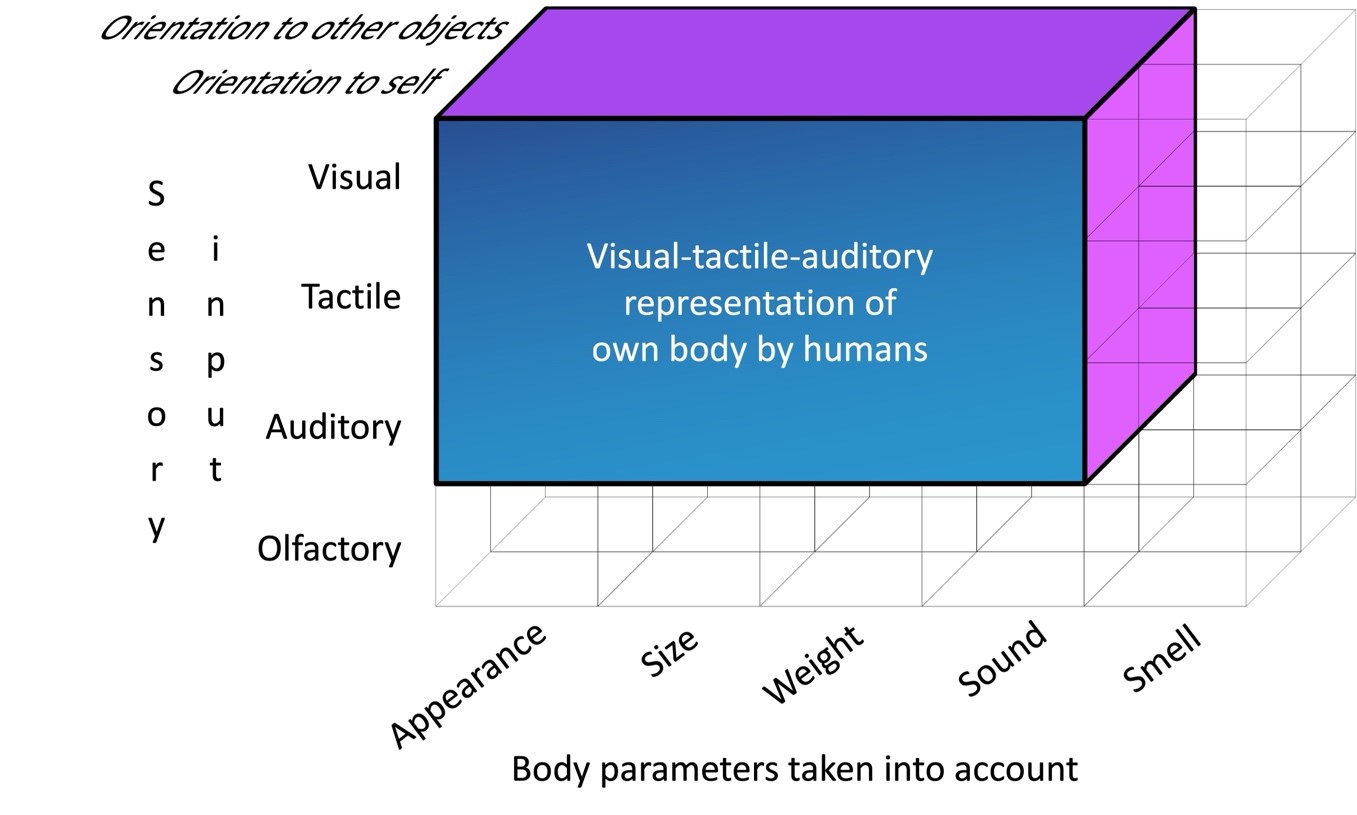
Figure 4. Modal organization of the visual-tactile-auditory module in humans (for explanation see the text).
The emergence of the visual-tactile-auditory module of self-representation could have been caused by the long-term evolution of the tool activity of apes and hominids (Toth & Schick, 2015). In addition, the evolution of the mind of representatives of the genus Homo was associated with the growth of cognitive fluidity, due to which previously isolated modules (like tools in a Swiss army knife (Cosmides & Tooby, 1994)) were connected to each other, forming larger highly integrative structures (Mithen, 1996; 2013; 2023). It is appropriate to note that the idea of cognitive fluidity widely circulated in recent years is consistent with the basic provisions of the Differentiation-Integration Approach (Chuprikova 2007, 2009). According to it, the development of any system begins with a homogeneous undifferentiated whole, then giving way to greater differentiation and autonomy of individual components, and later coming to a new integration.
Of course, this module of human self-representation is not the only one. In particular, the olfactory modality and the perception of one’s own body smell deserve special attention, even taking into account the evolutionary reduction of olfaction. Nevertheless, the functional load of the visual-tactile-auditory self-representation subsystem is extremely high, which probably makes it the leading one in terms of regulation of human behavior. In particular, this may explain certain anthropocentrism and the presentation of passing the MSR test as a sign of very high development of self-awareness (de Waal, 2019).
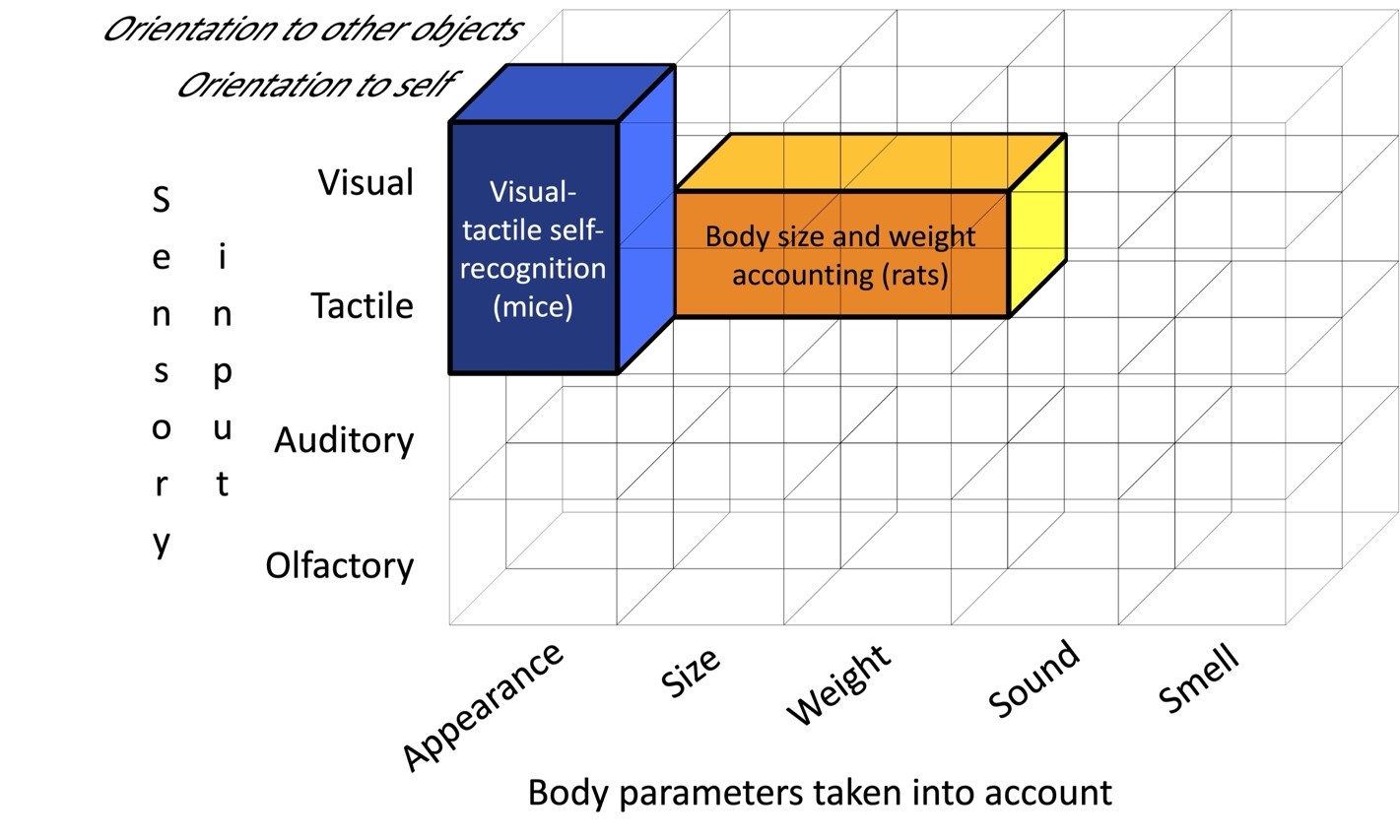
Figure 5. Modal organization of the visual-tactile-auditory module in humans (for explanation see the text).
Indeed, the research results suggest that animals have a larger number of more differentiated and functionally specialized modules for representing their own bodies. Considering rodents (mice and rats) as an example, we have the reason to describe two such modules (Fig. 5). First, the subsystem of tactile accounting of the size and weight of one’s own body when interacting with surrounding objects (Khvatov, Kharitonov, 2021; Krupa et al., 2001). Second, mice perform mirror-induced self-directed behavior, that is, they demonstrate signs of recognizing their own appearance, but only in the presence of tactile stimulation from marks applied to their bodies (Yokose et al., 2023).
It should be emphasized that due to the paucity of empirical material available to date, the above descriptions of self-representation modules are only plausible hypotheses that require further verification and clarification. Nevertheless, the proposed concept of the modular-modal organization of the representation of one’s own body, firstly, allows for a consistent interpretation of the existing data. In addition, it has an heuristic value in the sense that it provides grounds for putting forward and testing hypotheses about the specificity of the representation of one’s own body in previously unstudied animal species, taking into account the peculiarities of their ecology, physiology of sensory and effector systems, as well as behavior. For example, we can assume the presence of a module for representing various parameters of one’s own body (when focusing on external objects) based on the auditory modality in bats, taking into account their echolocation system (Müller, 2004) (Fig. 6).
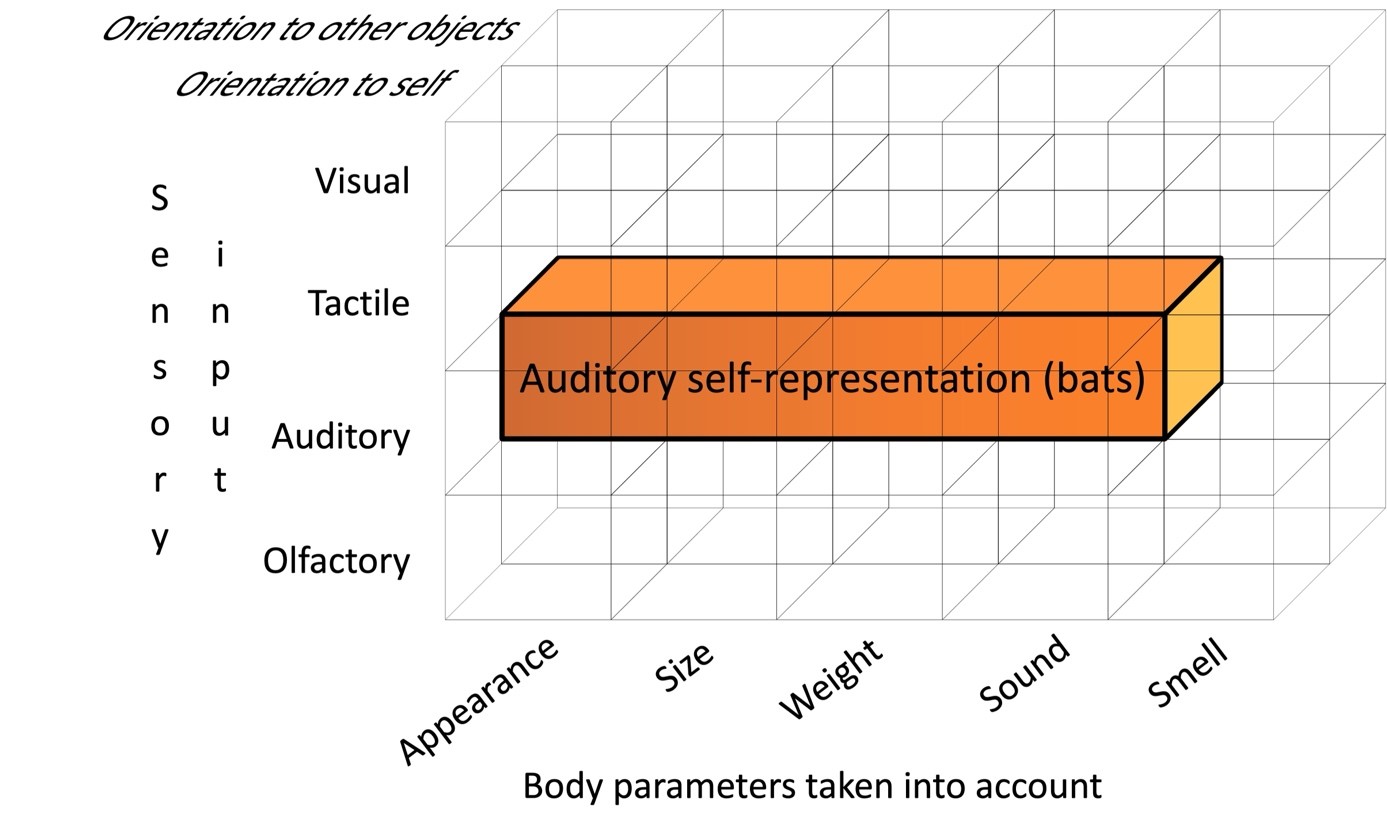
Figure 6. Self-representation module of bats based on auditory modality (see text for explanations).
Based on the above provisions of the model of modal-modular representation of one’s own body, below we formulate a number of requirements for experimental procedures that allow us to identify the specificity of functioning of self-representation subsystems in individual representatives of the animal kingdom.
One should take into account that the data on the features of animal representation of their bodies accumulated to date are rather scarce, and therefore many studies are exploratory. Accordingly, as noted earlier, the description of self-representation modules in many animals is to a major extent hypothetical. Nevertheless, it is necessary to clearly define the modality of the sensory input of self-representation and the modality of the body parameters taken into account, which are planned to be studied in representatives of a particular species, since the experimental procedure and setup directly depend on this. This should be done based on the available data on the features of ecology, morphophysiology of sensory and effector systems, as well as the behavior of the selected animal species. Even if studies of a certain self-representation module in the selected animal species have been previously conducted, understanding its modalities is necessary to avoid both methodological inaccuracies and interpretation errors.
The third modality deserves special attention, i.e. motivational orientation of the behavior regulation of which involves self-representation. Since the representation of one’s own body in the orientation of individual behavior to other objects is a basic type, it should first be established whether the animal is capable of considering as an obstacle certain parameters of its own body received by it through a certain sensory input. Accordingly, for this purpose it is necessary to use the “Body-as-obstacle task” methodology. Only after identifying this ability in a specific animal species is it advisable to study whether it demonstrates signs of a direct motivational orientation toward studying or modifying its own body. In this context, it is appropriate to recall the fair criticism of the MSR method in that many animals do not show such an interest in their own appearance as humans.
The Body-as-obstacle task technique involves animals demonstrating signs of their own body representation during interaction with surrounding objects to solve an external experimental task in which the goal is not their own body, but, for example, reaching a bait. The procedure for studying the features of animal’s representation of its body should include two stages. At the first stage, the animal, having encountered a certain experimental task, forms a certain behavioral algorithm for solving it in a series of trials. In addition, the experimental procedure should allow identifying predictors of the animal’s choice of a certain behavior in the current situation. At the second stage, the experimental setup is changed in such a way that the animal discovers the impossibility of satisfying its need with the previously developed behavioral algorithm. To modify its own behavior, the animal will need to take into account the physical parameters of its own body. In this case, the experimental plan should reveal precisely the animal’s ability to systematically choose (develop) the necessary behavior taking into account the features of its body, excluding the trial and error method.
To illustrate one possible realization of the above-mentioned theoretical considerations and requirements for experimental procedures of studying the characteristics of the representation of own body by animals, we present a method developed and repeatedly tested by us.
The subjects used were grey crows (Khvatov et al., 2021), domestic ferrets (Khvatov et al., 2023), and brown rats (Khvatov et al., 2024). Despite the great diversity, representatives of all species in their natural habitats are faced with tasks involving the penetration of various openings of their own body or its parts.

Figure 7. The experimental setup: 1—launch box; 2—finish box with bait; a, b, c—holes in the partition (Khvatov et al., 2023).
The experimental setup (see fig. 7) is a rectangular box divided into four parts by three partitions. The central partition has three holes, into which special inserts are placed, allowing the size and shape of the hole to be modified. The partitions at the edges have one large hole equipped with opaque impenetrable inserts, thereby forming the starting and finishing sections. The outer walls of the setup and the floor are made of light opaque organic glass, the partitions and inserts are made of opaque black organic glass. The specific sizes of the setup parts vary according to the species of test animals.
Experimental procedure comprises preliminary food deprivation. Before the test, the animal is placed in the starting box and the bait is placed in the finishing box. The experimenter then lifts the insert, allowing the subject to choose a hole to penetrate, after which the animal moves to the finishing box, where the bait is located. As soon as the subject takes the bait, the finishing compartment is closed with a partition, and the trial is over.
Two experimental series are conducted with animals of each species.
In the experimental trials of the first series, all three holes have different sizes (small, medium, and large), but all are permeable to the animals’ bodies. A hole of each size is located alternately on the left, in the center, and on the right 12 times, so the experimental session consists of 36 trials. The objective of the series is to identify predictors of the choice of a particular hole for penetration by the subject, i.e. its position in the partition (left, right, or center), as well as its size.
The purpose of the second experiment is to determine whether the animals can choose one passable hole out of the three, although the other two are larger but impassable. The experimental series consists of 72 trials. Two types of trials are used: test and background, alternating in such a way that two background trials followed one test. This is necessary to exclude the effect of learning.
In each of the 24 test trials, there is only one passable hole of a smaller area, and two impassable holes of a larger area. Two types of passable holes are used, i.e. round and square, and two types of impassable holes, i.e. rectangles with a greater height and rectangles with a greater width.
In each of the 48 background trials, two passable holes of a larger area and one impassable hole of a smaller area are used. Like in the test trials, the impassable holes are round and square, the passable holes are high narrow and low wide rectangles. The sizes of the holes also vary according to the species of animals.
In each trial, two dependent variable indices are registered, i.e. the first approach to the hole and the first attempt to pass through the hole. An “approach” is considered to be a situation when the distance between the tip of the muzzle or beak and the hole was no more than 1 cm. An “attempt” is considered to be a situation when the tip of the animal’s muzzle (or beak) penetrates the hole.
The conclusion about the animal’s ability to take into account the limits of its own body (that the animal plans its behavior using the subsystem for taking into account the size of its body) is made in the case of a reliable predominance of the first approaches followed by passage through a single penetrable hole in 24 test trials.
The results of the experiments on various species showed that three of the six crows (Khvatov et al. 2021), six ferrets (Khvatov et al., 2023) of the six tested and as well as six rats out of the six that took part in the experiments were able to take into account the boundaries of their own body when planning their behavior.
It should be noted that representatives of a number of other species, in particular amphibians (Khvatov & Zhelankin, 2018) and reptiles (Khvatov et al., 2019) may ignore the size of their bodies when entering holes without preliminary trial, but were able to learn this in a number of experimental trials. These facts substantiate an additional question about the heritability or acquisition of modules for representing one’s body during ontogenesis by different animals.
The repeated successful use of the method described above, in our opinion, not only evidences to its methodological validity, but also the validity of the theoretical model of modal-modular representation of one’s own body, of which this method is a realization.
Conclusions
1. Conclusion
The theory we propose models the phenomenon of animal representation of one’s own body in the form of a set of relatively autonomous modules, the specific qualities of which are revealed by means of considering three modalities: the type of parameters of one’s own body taken into account, the type of sensory input and the motivational orientation of the activity during which self-representation occurs.
As has been shown, this approach has a number of advantages. Firstly, the evolution of self-representation is now presented not as a monolithic linear process, but as a multitude of independent lines of development of forms of self-representation in different animal species in accordance with the unique features of their phylogeny, ecology and morphology. Such a model allows one to consistently generalize the accumulating empirical data on the features of body representation manifested by various animals. Secondly, based on the provisions of the proposed theoretical model, it is possible to formulate a number of requirements for the methodology of organization of the experiments aimed at studying the features of body representation in animals. In particular, one of such methods was developed by the authors and proved its effectiveness.
Funding: The research is supported by Russian Science Foundation, grant # 24-28-01049 (I.A.K) and Russian Ministry for Science and Education, project # 0138-2024-0012 (A.N.K.).
Credit author statement:
The authors declare no conflict of interest.
References
- Allen, M. R. (2007). Mirror self-recognition in a gorilla (Gorilla gorilla gorilla) Master thesis. Florida International University. doi:10.25148/etd.fi13101588
- Anderson, J. R. (1984). Monkeys with mirrors: Some questions for primate psychology. International Journal of Primatology, 5(1), 81–98. doi:10.1007/bf02735149
- Artoni, P., Chierici, M. L., Arnone, F., Cigarini, C., De Bernardis, E., Galeazzi, G. M., Minneci, D. G., Scita, F., Turrini, G., De Bernardis, M., & Pingani, L. (2021). Body perception treatment, a possible way to treat body image disturbance in eating disorders: a case-control efficacy study. Eating and Weight Disorders, 26(2), 499–514. doi:10.1007/s40519-020-00875-x
- Asendorpf, J. B., Warkentin, & V., & Baudonniere, P. M. (1996). Self-awareness and Other-awareness II: Mirror Self-recognition, social contingency awareness, and synchronic imitation. Developmental Psychology, 32(2), 313—321. doi:10.1037/0012-1649.32.2.313
- Barabanshchikov, V. A. (2002). Perception and event. Petersburg: Aleteya. [Barabanshchikov V. A. Vospriyatie i sobytie. SPb.: Aletejya, 2002].
- Barsalou, L. W. (2008). Grounded cognition. Annual Review of Psychology, 59, 617—645. doi: 10.1146/annurev.psych.59.103006.093639.
- Barsalou, L. W. (2010). Grounded cognition: past, present, and future. Topics in Cognitive Science, 2(4), 716—724. doi: 10.1111/j.1756-8765.2010.01115.x.
- Barth, J., Povinelli, D. J., & Cant, J.G.H. (2004). Bodily origins of Self. In D. Beike, J. Lampinen, & D. Behrend (Eds.), The Self and Memory. Psychology Press.
- Berti, A. & Frassinetti, F. (2000). When Far Becomes Near: Remapping of Space by Tool Use. of Cognitive Neuroscience, 12(3), 415—420. doi:10.1162/089892900562237
- Beulah, A. (1972). Mirror self-image reactions before age two. Developmental Psychobiology, 5(4), 297–305. doi:10.1002/dev.420050403
- Brownell, C. A., Zerwas, S., & Ramani, G. B. (2007). “So big”: the development of body self-awareness in toddlers. Child Development, 1426–1440. doi:10.1111/j.1467-8624.2007.01075.x
- Buniyaadi, A., Taufique, S. K. T., & Kumar, V. (2020). Self‐recognition in corvids: Evidence from the mirror‐mark test in Indian house crows (Corvus splendens). Journal of Ornithology, 161, 341–350. https://doi.org/10.1007/s10336-019-01730-2
- Burghardt, G. M., Partin, A. M., Pepper, H. E., Steele, J. M., Liske, S. M., Stokes, A. E., Lathan, A. N., Springer, C. M., & Jenkins, M. S. (2021). Chemically mediated self-recognition in sibling juvenile common gartersnakes (Thamnophis sirtalis) reared on same or different diets: evidence for a chemical mirror? Behaviour, 158, 1169–1191. doi:10.1163/1568539X-bja10131
- Carlson, T., Alvarez, G., Wu Daw-an Verstraten, F.A.J. (2010) Rapid Assimilation of External Objects Into the Body Schema. Psychological Science, 21(7), 1000—1005. doi:10.1177/0956797610371962
- Chuprikova, N. I. (2007). Mental development: The principle of differentiation. St. Petersburg: St. Petersburg. [Chuprikova N.I. Umstvennoe razvitie: Princip differenciacii. SPb.: Piter, 2007]
- Chuprikova, N. I. (2009). The universal differentiation and integration law of development as the basis of an interdisciplinary paradigm theory of development. Theory of development: A differentiation and integration paradigm. Moscow: Languages of Slavic Cultures, 7—16. [Chuprikova N.I. Vseobshchij universal’nyj differencionno-integracionnyj zakon razvitiya kak osnova mezhdisciplinarnoj paradigmal’noj teorii razvitiya. // Teoriya razvitiya: Differencionno-integracionnaya paradigma. M.: Yazyki slavyanskih kul’tur, 2009. S. 7–16]
- Coren, S. (2001). How to speak dog: mastering the art of dog-human communication. Atria Books.
- Coren, S. (2004). How dogs think: Understanding the Canine mind. New York: Free Press.
- Cosmides, L. & Tooby, J. (1994). Beyond intuition and instinct blindness: toward an evolutionarily rigorous cognitive science. Cognition, 50(1-3), 41-77. doi: 10.1016/0010-0277(94)90020-5. PMID: 8039372.
- Costantini, M., Frassinetti, F., Maini, M., Ambrosini, E., Gallese, V., & Sinigaglia, C. (2014). When a laser pen becomes a stick: remapping of space by tool-use observation in hemispatial neglect. Experimental Brain Research, 232(10), 3233—3241.
- Couchman, J. J. (2011). Self-agency in rhesus monkeys. Biology Letters, 8(1), 39–41. doi:10.1098/rsbl.2011.0536
- Dale, R. & Plotnik, J. M. (2017). Elephants know when their bodies are obstacles to success in a novel transfer task. Scientific Reports, 7. doi:10.1038/srep46309.
- de Vignemont, F. (2010). Body schema and body image – pros and cons. Neuropsychologia, 48(3), 669–680.
- de Waal, F. B. M. (2019). Fish, mirrors, and a gradualist perspective on self-awareness. PLoS Biology, 17(2). doi:10.1371/journal.pbio.3000112
- Delfour, F. & Marten, K. (2001). Mirror image processing in three marine mammal species: killer whales (Orcinus orca), false killer whales (Pseudorca crassidens) and California sea lions (Zalophus californianus). Behavioural Processes, 53(3), 181–190. doi:10.1016/s0376-6357(01)00134-6
- Denes, G. (1999). Disorders of body awareness and body knowledge. In Denes G, Pizzamiglio L (eds.). Handbook of Clinical and Experimental Neuropsychology. Psychology Press. pp. 497–506. doi:10.4324/9781315791272-28
- Di Vita, A., Boccia, M., Palermo, L., & Guariglia, C. (2016). To move or not to move, that is the question! Body schema and non-action oriented body representations: An fMRI meta- analytic study. Neuroscience & Biobehavioral Reviews, 68, 37–46.
- Dijkerman, H. C. & de Haan, E. (2007). Somatosensory processes subserving perception and action. Behavioral and Brain Sciences, 30(02),
- Dyson, B. J., Dunn, A. K., & Alain, C. (2010). Ventral and dorsal streams as modality-independent phenomena. Cognitive Neuroscience 1(1),
- Fabri, K. E. (2004). Fundamentals of zoopsychology (3rd ed.). Moscow: UMK “Psychology”. [Fabri, K. E. (2004). Osnovy zoopsihologii (3-e izd.). M.: UMK “Psihologiya”].
- Fein, S. & Spencer, S. J. (1997). Prejudice as self-image maintenance: Affirming the self through derogating others. Journal of Personality and Social Psychology. 73(1), 31–44. doi:10.1037/0022-3514.73.1.31
- Ferris, J. (2012). Self-awareness with a simple brain. Scientific American Mind, 23(5), 28–29.
- Filippova, G. G. (2012). Zoopsychology and comparative psychology. Moscow: Akademiya. [Filippova, G. G. (2012). Zoopsihologiya i sravnitel’naya psihologiya. M.: Akademiya].
- Fodor, J. A. (1983). The Modularity of Mind: An Essay on Faculty of Psychology, MIT.
- Fox, M. (1982). Are most animals “mindless automatons”? A reply to Gordon G. Gallup. American Journal of Primatology, 3, 341–343.
- Gallagher, S. (1986). Body image and body schema: A conceptual clarification. Journal of Mind and Behaviour, 7(4), 541–554.
- Gallup, G. G. Jr. (1970). Chimpanzees: Self recognition. Science, 167(3914), 86–87. doi:10.1126/science.167.3914.86
- Gallup, G. G. Jr. & Anderson, J. R. (2020). Self-recognition in animals: Where do we stand 50 years later? Lessons from cleaner wrasse and other species. Psychology of Consciousness: Theory, Research, and Practice, 7, 46–58.
- Garbarinia, F., Fossataroa, C., Bertia, A., Gindria, P., Romanod, D., Piaa, L., Gattaf, F., Maravitad, A., & Neppi-Modona, M. (2015). When your arm becomes mine: Pathological embodiment of alien limbs using tools modulates own body representation. Neuropsychologia, 70, 402-413. doi:10.1016/j.neuropsychologia.2014.11.008
- Gatti, R. C., Velichevskaya, A., Gottesman, B., & Davis, K. (2021). Grey wolf may show signs of self-awareness with the sniff test of self-recognition. Ethology Ecology & Evolution, 33(4), 444-467. doi:10.1080/03949370.2020.1846628
- Gibson, J. J. (1966). The Senses Considered as Perceptual Systems. Allen and Unwin, London.
- Gibson, J. J. (1979). The Ecological Approach to Visual Perception. Boston: Houghton Mifflin.
- Giglia, G., Pia, L., Folegatti, A., Puma, P., Fierro, B., Cosentino, G., Berti, A., & Brighina, F. (2015). Far Space Remapping by Tool Use: A rTMS Study Over the Right Posterior Parietal Cortex. Brain Stimulation, 8(4), 795-800. doi: https://doi.org/10.1016/j.brs.2015.01.412
- Gore, J. S. & Cross, S. E. (2011). Defining and Measuring Self-Concept Change. Psychological Studies, 56(1), 135–141. doi:10.1007/s12646-011-0067-0
- Gould, S. J. & Vrba, E. S. (1982). Exaptation—a Missing Term in the Science of Form. Paleobiology, 8(1), 4-15.
- Gozli, D. G. & Brown, L. E. (2011). Agency and Control for the Integration of a Virtual Tool into the Peripersonal Space. Perception, 40(11), 1309—1319.
- Hall, D. A. (2003). Auditory pathways: Are ‘what’ and ‘where’ appropriate? Current Biology, 13(10), R406–R408.
- Hant, G. T. (2004). About the nature of consciousness. From a cognitive, phenomenological and transpersonal point of view. Moscow: Publishing House of AST. [Hant, G. T. (2004). O prirode soznaniya. S kognitivnoj, fenomenologicheskoj i transpersonal’noj tochek zreniya. M.: Izd-vo AST].
- Hauser, M., Miller, C., Liu, K., & Gupta, R. (2001). Cotton-top tamarins (Saguinus oedipus) fail to show mirror-guided self-exploration. American Journal of Primatology, 137, 131–137. doi:10.1002/1098-2345(200103)53:3<131::AID-AJP4>3.0.CO;2-X
- Head, H. & Holmes, G. (1911). Sensory disturbances from cerebral lesions. Brain, 34(2–3), doi:10.1093/brain/34.2-3.102
- Hewitt, J. P. (2009). The Social Construction of Self-Esteem. In Lopez, Shane J.; Snyder, C.R. (eds.). Oxford Handbook of Positive Psychology. Oxford University Press. pp. 217–224.
- Hill, H. M., Webber, K., Kemery, A., Garcia, M., & Kuczaj, S. A. (2015). Can sea lions’ (Zalophus californianus) use mirrors to locate an object? International Journal of Comparative Psychology, doi:10.46867/ijcp.2015.28.00.08
- Horowitz, A. (2017). Smelling themselves: Dogs investigate their own odours longer when modified in an “olfactory mirror” test. Behavioural Processes, 143, 17-24. doi:10.1016/j.beproc.2017.08.001.
- Hotta, T., Komiyama, S., & Kohda, M. (2018). A social cichlid fish failed to pass the mark test. Animal Cognition, 21(1), 127–136. doi:10.1007/s10071-017-1146-y
- James, W. (1911). Psychology. St. Petersburg. [Dzhejms U. Psihologiya. SPb., 1911]
- Johnson-Frey, S. (2004). The neural bases of complex tool use in humans. Trends in Cognitive Sciences, 8(2), 71—78. doi:10.1016/j.tics.2003.12.002
- Kaplan, J. T., Aziz-Zadeh, L., Uddin, L. Q., & Iacoboni, M. (2008). The self across the senses: an fMRI study of self-face and self-voice recognition. Social Cognitive and Affective Neuroscience, 3(3), 218–223. doi:10.1093/scan/nsn014
- Kaplan, L. A., Wiegner, T. , Newbold, J. D., Ostrom, P. H., & Gandhi, H. (2008). Untangling the complex issue of dissolved organic carbon uptake: a stable isotope approach. Freshwater Biology, 53, 855-864. doi:101111/j.1365-2427.2007.01941x
- Keizer, A., Smeets, M. A., Dijkerman, H. C., Uzunbajakau, S. A., van Elburg, A., & Postma, A. (2013). Too fat to fit through the door: first evidence for disturbed body-scaled action in anorexia nervosa during locomotion. PLOS ONE, 8(5). doi:10.1371/journal.pone.0064602
- Khvatov, I. (2011). The concept of the origin of the psyche of A. N.Leontiev at the present stage of the development of science. Psychological Research, 4(15). https://doi.org/10.54359/ps.v4i15 .873 [Khvatov, I. (2011). Koncepciya proiskhozhdeniya psihiki A.N.Leont’eva na sovremennom etape razvitiya nauki. Psihologicheskie issledovaniya, 4(15). https://doi.org/10.54359/ps.v4i15.873].
- Khvatov, I.A.; Ganza, P.N.; Kharitonov, A.N.; Samuleeva, M.V. Wistar Male Rats (Rattus norvegicus domestica) Are Aware of Their Dimensions. Animals 2024, 14, 3384. https://doi.org/10.3390/ani14233384
- Khvatov, I. A., Kharitonov, A. N., & Sokolov, A. Y. (2021). Brown rats may learn awareness of their body weight when interacting with environmental objects. Cognitive Research, Artificial Intelligence and Neuroinformatics, Proceedings of the 9th International Conference on Cognitive Sciences, Intercognsci-2020/ Velichkovsky B. M., Balaban P. M., Ushakov V. L. Cham, Switzerland: Springer, 2021. Р.1358.
- Khvatov, I. A., Smirnova, A. A., Samuleeva, M. V., Ershov, E. V., Buinitskaya, S. D., & Kharitonov, A. N. (2021). Hooded crows (Corvus cornix) may be aware of their own body size. Frontiers in Psychology, 12. doi:10.3389/fpsyg.2021.769397.
- Khvatov, I. A., Sokolov, A. Y., & Kharitonov, A. N. (2019) Snakes elaphe radiata may acquire awareness of their body limits when trying to hide in a shelter. Behavioral Sciences, (9)67. doi:10.3390/bs9070067.
- Khvatov, I. A., Sokolov, A. Y., & Kharitonov, A. N. (2023). Ferrets (Mustela furo) are aware of their dimensions. Animals (Basel), 13(3), doi:10.3390/ani13030444.
- Khvatov, I. A., Zhelankin, R. V. (2018). Features of learning in toads Bufo viridis during behavior that requires taking into account the limits of their own bodies. Experimental Psychology, 11(4), 5-16. [Eksperimental’naâ psihologiâ (Russia)] doi: 10.17759/exppsy.2018110401.
- Kohda, M., Bshary, R., Kubo, N., Awata, S., Sowersby, W., Kawasaka, K., Kobayashi, T., & Sogawa, S. (2023). Cleaner fish recognize self in a mirror via self-face recognition like humans. Proceedings of the National Academy of Sciences, U S A, 120(7). doi: 10.1073/pnas.2208420120.
- Kohda, M., Hotta, T., Takeyama, T., Awata, S., Tanaka, H., & Asai, J.-y. (2019). If a fish can pass the mark test, what are the implications for consciousness and self-awareness testing in animals? PLoS Biology, 17(2). doi:10.1371/journal.pbio.3000021
- Krachun, С., Lurz, R., Mahovetz, L., & Hopkins, W. D. (2019). Mirror self-recognition and its relation – ship to social cognition in chimpanzees. Animal Cognition, 22(6), 1171–1183. doi:10.1007/s10071- 019-01309-7.
- Kraft, F. L., Forštová, T., Utku Urhan, A., Exnerová, A., & Brodin, A. (2017). No evidence for self-recognition in a small passerine, the great tit (Parus major) judged from the mark/mirror test. Animal Cognition, 20(6), 1049–1057. doi:10.1007/s10071-017-1121-7
- Krupa, D. J., Matell, M. S., Brisben, A. J., & Oliveira, L. M., Nicolelis, M. A. (2001). Behavioral properties of the trigeminal somatosensory system in rats performing whisker-dependent tactile discriminations. Journal of Neuroscience, 21(15), 5752-5763.
- Lei, Yanyu. (2023). Sociality and self-awareness in animals. Frontiers in Psychology, doi:10.3389/fpsyg.2022.1065638.
- Lenkei, R., Faragó, T., Kovács, D., Zsilák, В., & Pongrácz, Р. (2020). That dog won’t fit: body size awareness in dogs. Animal Cognition, 23, 337–350. doi:10.1007/s10071-019-01337-3
- Lenkei, R., Faragó, T., Zsilák, B., & Pongrácz, P. (2021). Dogs (Canis familiaris) recognize their own body as a physical obstacle. Scientific Reports, 11,
- Leontiev, A. N. (1981). Problems of mental development (4th ed.). Moscow: Publishing House of the Moscow University. [Leontiev, A. N. (1981). Problemy razvitiya psihiki (4-e izd.). M.: Izd-vo Moskovskogo universiteta].
- Leontiev, A. N. (2024). Lectures on general psychology. Moscow: Publishing House “Smysl”. [Leont’ev A. N. Lekcii po obshchej psihologii. Moskva, Smysl, 2024]
- A. N. (1975). Activity. Conscience. Personality. Moscow: Publishing House “Politizdat”. [Leontiev. A.N. Deyatel’nost’. Soznanie. Lichnost. M.: Politizdat, 1975].
- Levin, K. (2000). Field theory in the social sciences. [Teoriya polya v social’nyh naukah / Per. E. Surpina. — SPb.: Rech’, 2000].
- Lyon, P. (2015) The cognitive cell: bacterial behavior reconsidered. Frontiers in Microbiology, doi: 10.3389/fmicb.2015.00264
- Lyon, P., Keijzer, F., Arendt D., & Levin, M. (2021). Reframing cognition: getting down to biological basics. Philosophical Transactions of the Royal Society. B376: 20190750. https://doi.org/10.1098/rstb.2019.0750
- Ma, X., Jin, Y., Luo, B., Zhang, G., Wei, R., & Liu, D. (2015). Giant pandas failed to show mirror self-recognition. Animal Cognition, 18(3), 713–721. doi:10.1007/s10071-015-0838-4
- Maravita, A. & Iriki, A. (2004). Tools for the body (schema). Trends in Cognitive Sciences, 8 (2), 79–86. doi:10.1016/j.tics.2003.12.008
- Marfenin, N. N. (2016). A decentralized organism on the example of colonial hydroids. Interdisciplinary scientific and applied Journal “Biosphere”, 8(3), 315-337. [Marfenin N.N. Decentralizovannyj organizm na primere kolonial’nyh gidroidov // Mezhdisciplinarnyj nauchnyj i prikladnoj zhurnal «Biosfera», 2016, t. 8, No 3, S. 315-337].
- Markus, H. (1977). Self-schemata and processing information about the self. Journal of Personality and Social Psychology, 35(2), 63–78. doi:10.1037/0022-3514.35.2.63
- Mather, J. A. & Kuba, M. J. (2013). The cephalopod specialties: complex nervous system, learning, and cognition. Canadian Journal of Zoology, 91(6), 431–449. doi:10.1139/cjz-2013-0009
- Milner, A. D. & Goodale, M. A. (1995). The visual brain in action. Oxford University Press. 2d ed. (2006), pp. 157–181.
- Milner, A. D. & Goodale, M. A. (2008). Two visual systems re-viewed. Neuropsychologia, 46(3), 774 – 777.
- Mithen, S. J. (1996). The Prehistory of the Mind: A Search for the Origins of Art, Religion, and Science. London: Thames and Hudson, 1996.
- Mithen, S. J. (2013). The Cathedral Model for the Evolution of Human Cognition. Evolution of Mind, Brain, and Culture, edited by Gary Hatfield and Holly Pittman, Philadelphia: University of Pennsylvania Press, pp. 217-234. https://doi.org/10.9783/9781934536605.217
- Mithen, S.J. (2023). The evolutionary foundations of Neolithic thought: The invention of words, cognitive fluidity and objects as cognitive anchors. In L.E. Bennison-Chapman (ed.) Bookkeeping Without Writing. Early Administrative Technologies in Context. PIHANS CXXXIV. Netherlands Institute for the Near East.
- Moeller, B., Zoppke, H., & Frings, C. (2016). What a car does to your perception: Distance evaluations differ from within and outside of a car. Psychonomic Bulletin & Review, 23(3), 781-788. doi: 10.3758/s13423-015-0954-9.
- Moore, C., Mealiea, J., Garon, N., & Povinelli, D. J. (2007). The Development of Body Self- Awareness. Infancy, 11(2), 157–174.
- Müller, R. (2004). A numerical study of the role of the tragus in the big brown bat”. The Journal of the Acoustical Society of America. 116 (6): 3701–3712
- Norman, D. A. (2013). The design of everyday things (Revised and expanded editions ed.). Cambridge, MA London: The MIT Press.
- Paillard, J. (1999). Body Schema and Body Image-a Double Dissociation in deafferented patients. In G. Gantchev, S. Mori, & J. Massion (Eds.), Motor control today and tomorrow. Sofia, Bulgaria: Academic Publishing House.
- Pitron, V. & de Vignemont, F. (2017). Beyond differences between the body schema and the body image: insights from body hallucinations. Consciousness and Cognition, 53, 115–121. doi:10.1016/j.concog.2017.06.006
- Plotnik, J. M., de Waal, F.B.M., & Reiss, D. (2006). Self-recognition in an Asian elephant. Proceedings of the National Academy of Sciences, 103(45). doi:10.1073/pnas.0608062103
- Povinelli, D. J., Rulf, A. B., Landau, K. R., & Bierschwale, D. T. (1993). Self-recognition in chimpanzees (Pan troglodytes): distribution, ontogeny, and patterns of emergence. Journal of Comparative Psychology, 107(4), 347–372. doi:10.1037/0735-7036.107.4.347
- Prior, H., Schwarz, A., & Güntürkün, O. (2008). Mirror-induced behavior in the magpie (Pica pica): Evidence of self-recognition. PLOS Biology, 6(8). doi:10.1371/journal.pbio.0060202
- Rajala, A. Z., Reininger, K. R., Lancaster, K. M., & Populin, L. C. (2010). Rhesus monkeys (Macaca mulatta) do recognize themselves in the mirror: Implications for the evolution of self-recognition. PLOS ONE, 5(9). doi:10.1371/journal.pone.0012865
- Reiss, D. & Marino, L. (2001). Mirror self-recognition in the bottlenose dolphin: A case of cognitive convergence. Proceedings of the National Academy of Sciences, 98(10), 5937–5942. doi:10.1073/pnas.101086398
- Ritchie, J. B. & Carlson, T. A. (2013). Tool Integration and Dynamic Touch. Psychological Science, 4(6), 1066-8. doi:10.1177/0956797612459768
- Robinson, T. (2023). Mirror self-recognition in ghost crab (Ocypode quadrata). Animal Cognition, 26(5), 1539–1549. doi:10.1007/s10071-023-01800-2. PMID 37314594
- Rochat, P. (2003). Five levels of self-awareness as they unfold early in life. Consciousness and Cognition 12(4), 717–731. doi:10.1016/s1053-8100(03)00081-3
- Rogers, K. R. (1994). A look at psychotherapy. The formation of man. Moscow: Publishing group “Progress”, “Univers”, 1994. [Rodzhers K. R. Vzglyad na psihoterapiyu. Stanovlenie cheloveka. — M.: Izdatel’skaya gruppa «Progress», «Univers», 1994]
- Roma, P., Silberberg, A., Huntsberry, M., Christensen, C., Ruggiero, A., & Suomi, S. (2007). Mark tests for mirror self-recognition in Capuchin monkeys (Cebus apella) trained to touch marks. American Journal of Primatology, 69(9), 989–1000. doi:10.1002/ajp.20404
- Romanski, L. M., Tian, B., Fritz, J., Mishkin, M., Goldman-Rakic, P. S., & Rauschecker, J. P. (1999). Dual streams of auditory afferents target multiple domains in theprimate prefrontal cortex. Nature Neuroscience 2(12), 1131–1136.
- Schilder, Paul.The image and appearance of the human body. (1935). London.
- Schiffner, I., Vo, H. D., Bhagavatula, P. S., & Srinivasan, M. V. (2014). Minding the gap: in-flight body awareness in birds. Frontiers in Zoology, 11, 64. doi:10.1186/s12983-014-0064-y
- Shaffer, V. A. & Renner, M. J. (2000). Black-and-White Colobus Monkeys (Colobus guereza) do not Show Mirror Self-Recognition. International Journal of Comparative Psychology, 13(3). doi:10.46867/C4H01J
- Shannon, C. E. (1948). A mathematical theory of communication. Bell System Technical Journal, 27(3), 379–423. doi:10.1002/j.1538-7305.1948.tb01338.x.
- Stolin, V. V. (1983). Self-awareness of personality.: Publishing House of the Moscow University. [Stolin, V. V. (1983). Samosoznanie lichnosti. M.: Izd-vo Moskovskogo universiteta].
- Suarez, S. D & Gallup, G. G. (1981). Self-recognition in chimpanzees and orangutans, but not gorillas. Journal of Human Evolution, 10(2), 175–188. doi:10.1016/s0047-2484(81)80016-4
- Szabo, B. & Ringler, E. (2023). Geckos differentiate self from other using both skin and faecal chemicals: evidence towards self-recognition? Animal Cognition, 26, 1-9. doi:10.1007/s10071-023-01751-8.
- Tabrik, S., Behroozi, M., Schlaffke, L., Heba, S., Lenz, M., Lissek, S., Güntürkün, O., Dinse, H.R., & Tegenthoff, M. (2021). Visual and tactile sensory systems share common features in object recognition. eNeuro, 8(5). doi: 10.1523/ENEURO.0101-21.2021. PMID: 34544756; PMCID: PMC8493885.
- The Cambridge Declaration on Consciousness (Archive) 7 July 2012. Written by Philip Low and edited by Jaak Panksepp, Diana Reiss, David Edelman, Bruno Van Swinderen, Philip Low and Christof Koch. University of Cambridge.
- The New York Declaration on Animal Consciousness New York University, 19 April 2024. URL: https://sites.google.com/nyu.edu/nydeclaration/declaration
- Thompson, E. (2010). Mind in life: Biology, phenomenology, and the sciences of mind. Harvard University Press.
- Toth, N. & Schick, K. (2015). Chapter 14 – Evolution of Tool Use. In Basics in Human Editor(s): Michael P. Muehlenbein, Academic Press, 2015, Pages 193-208, https://doi.org/10.1016/B978-0-12-802652-6.00014-1.
- Turner, J. & Oakes, P. (1986). The significance of the social identity concept for social psychology with reference to individualism, interactionism and social influence. British Journal of Social Psychology, 25(3), 237–252. doi:10.1111/j.2044-8309.1986.tb00732.x
- Ukhtomsky, A. A. (1927). Physiology of the motor apparatus. Moscow: “Practical Medicine”. [Uhtomskij A. A. (1927). Fiziologiya dvigatel’nogo apparata. M.: «Prakticheskaya medicina»].
- Varela, F. J., Thompson, E. T., & Rosch, E. (1991). The embodied mind: Cognitive science and human experience. Cambridge, MA: The MIT Press.
- Walraven, V., van Elsacker, L., & Verheyen, R. (1995). Reactions of a group of pygmy chimpanzees (Pan paniscus) to their mirror images: evidence of self-recognition. Primates, 36(1), 145–150. doi:10.1007/bf02381922
- Warren, J. E., Wise, R. J., & Warren, J. D. (2005). Sounds Do-Able: Auditory-motor transformations and the posterior temporal plane. Trends in Neurosciences, 28(12), 636–643.
- Warren, Jr. W. H. & Whang, S. (1987) Visual guidance of walking through apertures: body-scaled information for affordances. Journal of experimental psychology: human perception and performance, 13(3),
- Wilson, M. (2002). Six views of embodied cognition. Psychonomic Bulletin and Review, 9(4), 625–636. doi:10.3758/BF03196322
- Yokose, J., Marks, W.D., & Kitamura, T. (2023). Visuotactile integration facilitates mirror-induced self-directed behavior through activation of hippocampal neuronal ensembles in mice. Neuron, 112(2), 306-318. doi: 10.1016/j.neuron.2023.10.022. Epub 2023 Dec 5. PMID: 38056456.
- Zatorre, R. B., Marc A., & Pierre B. P. (2002). Where is ‘where’ in the human auditory cortex? Nature neuroscience, 5, 905-9.
- Zinchenko, V. P. (1999). The problem of external and internal formation of the image of oneself and the world as the realization of consciousness. The world of psychology, 1, 97 – 104. [Zinchenko V. P. Problema vneshnego i vnutrennego stanovleniya obraza sebya i mira kak realizaciya soznaniya. Mir psihologii, 1, 97 – 104]
- Zorina, Z. A., Poletaeva, I. I. (2003). Zoopsychology: elementary thinking of animals: A textbook for universities. M.: Aspect Press. [Zorina, Z. A., Poletaeva, I. I. (2003). Zoopsihologiya: elementarnoe myshlenie zhivotnyh: Uchebnoe posobie dlya vuzov. M.: Aspekt Press]
Comments (0)
Studies in animal self-awareness is a most promisimg direction in the field of evolutionary and comparative psychology. Different experimental methods were developed and tested provoding a vast amount of contradictory data. These data gave rise to a lot of difficulties and discordance in theoretical justification of the phenomenon of self-representation. We propose a model of body representation in animals, within the framework of which the self-representation is viewed not as a sort of monolithic abstract knowledge of an individual about the self, but as a set of modality-specific subsystems. The purpose of such dedicated systems is to process various physical characteristics of the individual’s body as related to solving specific behavioral problems. Based on the provisions of this approach, a number of requirements are formulated for the methods aimed at studying self-awareness in animals. Also, using the ideas of the Differentiation-Integration approach, the main trends in the evolution of self-representation may be identified.
Представление о Телесном Осознании в Терминах Модальности и Модульности
Хватов И.А. а, Харитонов А.Н. б*
аНаучно-образовательный центр Биопсихологических исследований Московского института психоанализа, 121170 Москва, Россия ORCID iD: 0000-0002-6939-244X
бИнститут психологии Российской академии наук, 129366 Москва, Россия, ORCID iD: 0000-0002-4801-9937
Аннотация: За последнюю четверть века изучение Self-awareness стало одним из наиболее бурно-развивающихся научных направлений в области Animal cognition. Было разработано и апробировано множество различных экспериментальных методик, накоплен обширный объем противоречивых данных, что породило немало трудностей в теоретическом осмыслении феномена саморепрезентации у животных. Мы предлагаем оригинальную теорию репрезентации собственного тела животными, в рамках которой саморепрезентации представляется не в качестве единого абстрактного знания индивида о себе, но в виде множество модально специфических модулей, учитывающих различные физические характеристики тела индивида для решения частных поведенческих задач. На основе положений развиваемой теории формулируется ряд требований к методикам, нацеленным на изучение Self-awareness у животных. С привлечением дифференционно-интеграционного подхода выделяются основные тенденции эволюции саморепрезентации.
Ключевые слова: самопрезентация, представление о собственном теле, модальность самопрезентации, модули самопрезентации, самосознание, осознание тела, зеркальное самопознание, дифференционно-интеграционный подход.
Информация об авторах
Хватов Иван Александрович, кандидат психологических наук, заведующий кафедрой общей психологии, руководитель Научно-образовательного центра бмопсихологических исследований, НОЧУ ВО Московский институт психоанализа, г. Москва, Российская Федерация, ORCID: https://orcid.org/0000-0002-6939-244X, e-mail: ittkrot1@gmail.com
Харитонов Александр Николаевич, кандидат психологических наук, старший научный сотрудник, Инсттитут психологии РАН, г. Москва, Российская Федерация, ORCID: https://orcid.org/0000-0002-4801-9937, e-mail: ankhome47@list.ru
Self-representation is a mental representation of an individual’s own qualities, which is necessary for regulating their behavior and interaction with the environment. This concept denotes an extremely wide range of diverse phenomena: self-concept (Rogers, 1994; Gore, Cross, 2011); self-image (Fein, Spencer, 1997); self-schema (Markus, 1977), self-esteem (Hewitt, 2009), social identity (Turner, Oakes, 1986), etc. In humans, most of these types and/or forms of self-representation are culturally and socially determined (Stolin, 1983).
Which form of self-representation is the phylo- and ontogenetically earliest? In answering this question, one may be guided by the fundamental principle: the initial mental images in terms of development contain a minimum of abstractions, being direct representations of specific objects, i.e. material bodies (Zorina, Poletaeva, 2003). Accordingly, the earliest (most probably, the starting point), both in phylo- and ontogenesis, should be considered the mental representation of one’s own body, an individual’s idea of the physical qualities of own body necessary to regulate his behavior and interaction with the environment (Khvatov et al., 2023). This phenomenon has been discussed for a long time. W. James called it the “physical self” (1911), and V.V. Stolin (1983) proposed the idea of “taking oneself into account”. Nevertheless, the theoretical development of such a seemingly simple concepts is still far from complete.
Currently, two forms of representation of one’s own body are distinguished, i.e. body schema and body image. Body schema is an unconscious (implicit), sensorimotor representation that arises directly in the course of behavior, which is the basis for constructing an egocentric coordinate system (Head & Holmes, 1911; Keizer et al., 2013). A significant part of the research in the body schema and the possibilities of its modification is carried out on people (Johnson-Frey, 2004). The body schema is highly differentiated and flexible, integrating into its structure external objects that are in physical contact with the individual, for example, the tools used by him (Carlson et al., 2010; Carlson, 2013; Costantini et al., 2014; Garbarinia et al., 2015; Gozli & Brown, 2011; Ritchie & Moeller et al., 2016). In accordance with the change in the body scheme, there is also a modification of the subjective perception of the surrounding reality, i.e. the egocentric coordinate system (Barabanschikov, 2002; Berti & Frassinetti, 2000; Giglia et al., 2015; Costantini et al., 2014).
Body image is a conscious (explicit) representation of one’s own body characteristics (e.g., one’s own appearance), which serves as the basis for constructing an allocentric coordinate system (Schilder, 1935). This phenomenon is often studied clinically. Distortions of body image are observed in various psychiatric and neurological disorders (Denes, 1999; Artoni et al., 2021).
Apart from psychology, distinction between body schema and body image has been the basis of much work in neuropsychology, cognitive neuroscience and philosophy (de Vignemont, 2010; Dijkerman & de Haan, 2007; Di Vita et al., 2016; Gallagher, 1986; Paillard, 1999). The body schema functions directly in the process of behavior, whereas body image concerns the abstract imagination of a complex of one’s own physical properties. This distinction is consistent with the general functional hypothesis according to which perception and action require different transformations of sensory signals, follow different rules and are thus serviced by different cortical and subcortical pathways. This has been shown for visual and auditory signaling (Dyson, Dunn, & Alain, 2010; Hall, 2003; Milner & Goodale, 1995; Milner & Goodale, 2008; Romanski et al., 1999; Warren, Wise, & Warren, 2005; Zatorre et al., 2002) as well as for somatosensory systems (Dijkerman, de Haan, 2007). However, in most everyday situations, the “perceived body” does not confront the “acting body”. In this regard, it has been argued that the discussed difference between body schema and body image is putative and these two phenomena act as different functional mannifestations of individual’s representation of one’s own body (Pitron & de Vignemont, 2017).
A completely different narrative emerged in the fields of evolutionary and comparative psychology that study the phylogeny and species-specific diversity of forms of body representation in animals. Firstly, in this case the authors do not focus on the differences between the body scheme and the body image. There are only isolated studies of the body scheme in animals in the field of neuroscience (Maravita, & Iriki, 2004). This is probably due to the fact that most experiments conducted on animals we discuss below are behavioral, that is, directly related to the body scheme, and therefore it is not possible to isolate the body image only. Secondly, to designate self-representation, as applied to animals, in the English-language literature the researchers mostly use the term “self-awareness”, that is the ability of an individual to perceive himself (primarily the body) separately from the characteristics of the external world and/or other individuals (Ferris, 2012).
We have deliberately avoided using the term “self-awareness” so far, since there are currently disagreements in cognitive science, which is dominant throughout the world. Among the proponents of the Cultural Activity Approach which is widely spread in the post-Soviet space there is also much controversy regarding its applicability to animals.
In the English-language literature, authors operate with the concept of “self-awareness” rather loosely identifying its features not only in humans, but also in a huge number of different species (de Waal, 2019; Gallup & Anderson, 2020; Hunt, 2004). It should be mentioned here that promotion of “animal consciousness” is also driven by the interests of bioethics (The Cambridge Declaration on Consciousness, 2012; The New York Declaration on Animal Consciousness, 2024), which has no direct relation to science. So we will allow ourselves to leave this aspect without comment.
Unlike in cognitive psychology, the Cultural Activity Approach has a clearer understanding of consciousness, considering it as a level of mental evolution specific to humans (Filippova, 2012). In this regard, Russian comparative psychology is not so much concentrated on animal consciousness, as on the phylogenetic prerequisites of human consciousness (Zorina & Poletaeva, 2003). This topic has been developed in detail in the works of Russian psychologists (Fabri, 2004; Filippova, 2012; Leontiev, 1981). A.N. Leontiev defines consciousness as a reflection of objective reality in which its objective stable properties are distinguished regardless of the subject’s attitude towards it. The impetus for the formation of consciousness is placed on labor as a specific collective activity, involving the division of functions between several participants, which, in turn, determines the emergence of language generated by human relationships with other people that develop in the process of labor (Leontiev, 1981). Consequently, self-awareness is considered one of the key components of consciousness as the highest level of phylo- and ontogenetic development of the mind inherent in humans and distinguishing them from other animal species (Filippova, 2012; Leontiev, 1981), even when it comes to such a relatively simple phenomenon as awareness of one’s own body.
In our opinion, the current terminological dispute is due to the differences in the theoretical foundations of the two approaches. Whereas cognitive psychology is predominantly based on positivism and neo-positivism, inherited from behaviorism, the Cultural Activity Approach is based on the sociologically oriented ideology of dialectical materialism. Without going into further details of methodological discussions, we should admit that the question of emergence of the representation of one’s own body (as the basis for other types and forms of self-representation) in the course of phylogenesis remains open and attracts much attention.
Figure 1. Two models of the evolution of self-representation (English: “self-awareness”): A – binary, B – gradualistic (see text for explanations) (from de Waal, 2019).
Currently there are two most widely adopted models that are relevant to explain the emergence of representation of one’s own body in animals in the process of mental evolution: binary and gradualistic (Fig. 1).
- The binary model postulates sudden emergence of self-representation, the presence of which can be judged by passing the mirror self-recognition (MSR) test. In 1970, Gordon Gallup Jr. experimentally investigated the possibility of self-recognition in a mirror in two naive males and two naive females of wild chimpanzees (Pan troglodytes). Each chimpanzee was first placed in an experimental room for two days. Then a full-length mirror was placed in the room for 80 hours. Various behavioral patterns were then recorded during the interaction of the monkeys with the mirror. Initially, the chimpanzees made threatening gestures towards their own reflections, supposedly seeing them as a threat in return. By the end of the experiment, the chimpanzees used their own reflections to guide self-directed behaviors, such as grooming parts of their body not previously observed without a mirror, nose picking, and the like. Gallup extended the study by manipulating the chimpanzees’ appearance and observing their reactions to the mirror reflection (Gallup, 1970). Gallup lightly anesthetized the chimpanzees and then applied an alcohol-soluble dye to the edge of the eyebrow and the upper half of the opposite ear. When the dye dried, it had virtually no olfactory or tactile properties. Gallup then returned the chimpanzees to the cage (with the mirror removed) and they regained full consciousness. He then recorded the frequency with which the chimpanzees spontaneously touched the marked areas of their skin. After 30 minutes, the mirror was reintroduced into the room and the frequency of touching the marked areas was again registered. The frequency of touching the marked areas of the body was four to ten times higher in the presence of a mirror than in its absence.
Based on the results obtained, it was concluded that the chimpanzees remembered their appearance and noticed changes in it, and also understood that the image in the mirror was equivalent to their own body.
To date, the ability to recognize oneself in a mirror has been established in humans (starting from the age of two years) (Beulah, 1972), great apes (Allen, 2007; Povinelli et al., 1993; Suarez & Gallup, 1981; Walraven et al., 1995), dolphins (Reiss & Marino, 2001), killer whales (Delfour & Marten, 2001), Asian elephants (Plotnik et al., 2006), Eurasian magpies (Prior et al., 2008), Indian house crows (Buniyaadi et al., 2020), cleaner wrasses (Kohda et al., 2019; 2023), and Atlantic crabs (Robinson, 2023). On the other hand, many animals failed the MSR test, i.e. sea lions (Hill et al., 2015), giant pandas (Ma et al., 2015), dogs (Coren, 2001), monkeys (Hauser et al., 2001; Ma et al., 2015; Rajala et al., 2010; Roma et al., 2007; Shaffer, & Renner, 2000), grey parrots (Hill et al., 2015), great tits (Kraft et al., 2017), cichlid fish (Hotta et al., 2018), and octopuses (Mather & Kuba, 2013).
- The gradualistic model assumes that all species have self-awareness, as it is necessary for survival. Rochart identifies 5 levels of self-representation development in human ontogenesis (Rochat, 2003). Later Frans de Waal extrapolated this concept to phylogenesis (de Waal, 2019). From this point of view, representation of one’s own body can be imagined as an onion, i.e. the phenomenon is multifaceted, it becomes more complex and supplemented as the mind evolves. However, the highest developmental level of self-awareness is associated with passing the MSR test.
Thus, the above models, in one way or another, both assume that the highest level of development of the representation of one’s own body is expressed in the ability to recognize oneself in a mirror or in a photo.
One should note that there is much criticism of the MSR approach. Firstly, it cannot be applied to animals whose visual modality is not the leading one (most mammals) (Coren, 2004). Secondly, both animals and humans may not show interest in marks on their own bodies so far as they may not recognize this mark as something abnormal (Fox, 1982; Asendorpf et al., 1996). Thirdly, some animals may feel uncomfortable looking at “the other” in the mirror, since this provokes aggression (Anderson, 1984; Couchman, 2011). Fourthly, vertical reflective surfaces are almost never found in nature (Lei, Yanyu, 2023).
A persistent idea widely spread in scientific circles is that the ability to recognize oneself in a mirror, firstly, is an expression of complex high-level cognitive processes, and secondly, indicates the inclusion of the subject into flexible social connections and predisposition to understand the intentions of conspecifics, or “theory of mind” (Krachun et al., 2019; Lei, Yanyu, 2023). Meanwhile, the ability to pass the appropriate test is demonstrated by those animals whose mental regulation of behavior is extremely simple (however not primitive), i.e. fish (Kohda et al., 2019; 2023) and crabs (Robinson, 2023).
In summary, it should be concluded that at present the field of evolutionary and comparative studies of representation of own body in animals is in crisis signaled by the impossibility of consistently explaining the genesis and evolution of this phenomenon, as well as its species diversity. This crisis is caused by a number of both theoretical and methodological contradictions.
Below we propose an alternative model of the genesis and evolution of the representation of one’s own body by animals, which, in our opinion, can provide a basis for a consistent integration of the diverse empirical data in this area, as well as propose an efficient method for studying the phenomenon under discussion.
2. A model of modal-modular representation of one’s own body
The model of representation of one’s own body by animals that we propose is based on two provisions.
- In the process of mental regulation of behavior any individual must take into account the physical characteristics of the body (boundaries, volume, mass) and correlate them with the physical characteristics of external objects. In other words, animals need to “take themselves into account” (Stolin, 1983). This thesis is well substantiated by the concept of “embodied cognition” (Varela et al., 1991), which is being rapidly developed in modern cognitive psychology. The basic tenet of this concept is that the mind matures, exists and develops as a result of the interaction of the integral physical body (not just the brain or nervous system) with physical objects in the external environment, including social contacts. In particular, this means that cognition is rooted in the environment, it unfolds in the context of the surrounding world. Even autonomous (separated from the environment) cognition is based on the individual’s bodily experience (Wilson, 2002).
To develop an original model of self-representation based on the above provisions of Embodied Cognition Approach, we will need to clarify the most basic categories.
The mind here is considered as an information based process that regulates the behavioral interaction of an individual with the diversity of objects in the environment. In turn, behavior is a form of life activity of an individual (a unitary organism), providing adaptation by changing spatial relationship with objects of the environment by means of locomotor and manipulatory acts. In this context, a unitary organism as integral and indivisible entity (Marfenin, 2016) is opposed to a modular, decentralized one. Accordingly, many animals (but for sessile or parasitic), as well as heterotrophic protists, belong to unitary creatures: the mind is necessary for them to regulate their own behavior. It is reasonable to note that in the course of the discussion we inevitably come into contact with the problem of the genesis of the mind. However, a detailed analysis of this topic is not the purpose of this article, although it certainly deserves rethinking in the light of modern facts from the field of biology (Khvatov, 2011; Lyon, 2015; Lyon et al., 2021).
It is important to emphasize that the definition of the mind presented above is not radically new. Firstly, it is based on the traditional understanding of information as a process of removing uncertainty (Shannon, 1948). Secondly, this definition is entirely consistent with the classical Soviet psychology consideration of the mind through the prism of its involvement in the process of regulation of activity and behavior.
Accordingly, for the behavioral satisfaction of its own need, the animal each time creates subjective representations of a variety of external objects, as well as a representation of itself. Thus, the mind is located “on the edge” of two objective realities, external and internal, the reality of one’s own organism (Zinchenko, 1999). By constructing and synthesizing these representations, the individual builds an image of a holistic situation, the event (joint existence) (Barabanshchikov, 2002), thus providing a basis for behavioral planning. Kurt Lewin called this phenomenon the Psychological Field of Life Space (Lewin, 2000). For the possibility of behavioral satisfaction of an actual need, the synthetic structure of the image of the event must be harmonious in the sense of a complementary relationship between the representations of one’s own body and the surrounding objects. As a result, the planned set of locomotor and manipulatory acts is embodied in accordance with the physical limitations and affordances (Gibson, 1966; Norman, 2013) of both the individual’s body and the characteristics of current situation.
The problem arises in a situation when a behavioral act cannot be realized due to the presence of limitations of one’s own body and/or material environment. In such a case, the individual either discovers this circumstance at the planning stage, or makes a mistake, trying unsuccessfully to perform the behavioral act.
Then one of three possible scenarios is possible:
- an individual’s refusal to satisfy his needs behaviorally;
- an individual’s reorganization of the mental image of an event, taking into account the existing physical limitations of one’s own body and surrounding objects, in order to plan a new behavioral act;
- a systematic physical modification of one’s own body, surrounding objects, or both, in order to give them the required qualities for the possibility of implementing the original behavioral act.
The latter two options provide the basis for planning various psychological experiments to study the peculiarities of animal thinking (Zorina, Poletaeva, 2003; Fabri, 2004). As an example of the second option, we can cite the classic experiment to discover the detour (Fabri, 2004). To illustrate the third option, it is appropriate to mention Wolfgang Kohler’s experiments with chimpanzees. In this case, by constructing a tower of boxes, the chimpanzee modified the environment and, by extending his arm with a stick, he modified his own body.
It follows from the above that in any ethological experiment, in the process of problem solving when planning its behavior, an animal uses the representation of its own body in some way. The problem is that, for a researcher building a model of the animal’s mental processes on the basis of the obtained empirical data, it is often extremely difficult to isolate the self-representation, i.e. the representation of the parameters of animal’s own body, and separate it from the representation of the parameters of other surrounding objects. In other words, changing an independent variable we often cannot say for sure whether the tested animal takes into account its body or other objects when it changes its behavior. A better assumption would be it takes into account both, as its body and the external world in subjective representation do not exist in isolation from each other. This circumstance should be considered when designing experiments aimed at identifying the features of the representation of one’s own body by animals of certain species.
2. The second assumption concerns the structural organization of the representation of one’s own body. Here one may rely on the methodology of Grounded Cognition (Barsalou, 2008; 2010). Cognitive psychology traditionally represents cognition as a sort of computation occurring in modular systems that process abstract and amodal content. According to grounded cognition theory, cognition is based on modality-specific and bodily-conditioned processes. Modality is understood not as a sensory quality, but as the specificity of individual’s behavior within certain conditions when solving a specific problem. Accordingly, grounded cognition appears to be associated with the ideas of enactivism (Thompson, 2010), which proposes to consider a mental image not as a simple copy (representation) of objective reality, but as a system continuously constructed by an individual in the process of his behavioral activity, and therefore dependent on specificity of the latter. In turn, enactivism, which emerged in the 90ies of the past century, was in a way anticipated by the basic provisions of the Theory of Activity (Leontiev, 1975).
Bringing the above ideas into the framework of the model we are constructing suggests that the representation of one’s own body by an individual is not a monolithic system of storage of abstract information about all sorts of characteristics of oneself. We propose the self-representation is a set of subsystems modally specialized in relation to the individual’s mental regulation of his behavior under specific material conditions that require consideration of clearly defined parameters of his body.
Figure 2. Three modalities of representation of one’s own body (see text for explanations)
To further discuss the model, we introduce and analyze two concepts: the modality of self-representation and the module of self-representation. The modality of self-representation is a particular dimension of the representation of one’s own body by an individual arising in accordance with the specific features of behavioral activity and the ecological context within which this representation is a simple copy (representation) of objective reality, but as a system continuously constructed by an individual in the process embodied. We distinguish three modalities: sensory input, the parameters of one’s own body taken into account, and the motivational orientation of behavior within which self-representation functions (Fig. 2).
Sensory input is a specific quality of sensory data used to construct self-representation, something that A.N. Leontiev called “sensory fabric” (Leontiev, 2024). Here we distinguish four types of sensory inputs: visual, tactile, auditory and olfactory (Fig. 2) engaged in self-representation. It is important to note that this typology serves only as a theoretical starting point so far as it is possible to distinguish several more variants of sensory inputs in different animal species (e.g., electroreception, magnetoreception, etc.).
By the parameters of one’s own body we mean specific manifestations of the individual’s body taken into account by an individual when constructing a self-representation in the context of current behavior. To begin with, it is necessary to distinguish five types of such parameters: one’s own appearance, one’s own body size, one’s own body weight, one’s own body sound (both voice and other sounds that occur during locomotor and/or manipulative acts), and one’s own body odor. For example, when choosing a hole of a suitable size for penetration, an animal must take into account the size (boundaries) of its body, and when determining the strength of the support for passage its weight. At the same time, one should not confuse sensory input with the parameters of one’s own body taken into account. Naturally, under certain conditions they may coincide. Alternatively, looking at one’s own reflection in the mirror, we construct a representation of one’s own appearance based on the visual modality. On the other hand, a person, determining the penetrability of a certain opening (say, a doorway) with his own body and, accordingly, taking into account the size of the latter, also relies on visual data (Warren & Whang, 1987). A rat, due to weak development of its vision, solving a similar problem, is primarily guided by tactile sensations from its vibrissae (Krupa et al., 2001).
The third modality, motivational orientation, is subdivided into only two types and is determined according to whether the individual’s behavior is oriented toward self-study and/or self-modification (e.g., cleaning one’s body) or not. We shall discuss this modality in more detail, since its types affect two main methodological approaches currently used to study self-representation in animals.
If the subject is focused on self-study and/or self-modification (Fig. 2, blue blocks), we assume that his or her own body is a goal. The ability to represent one’s body for the purpose of self-modification is studied using the MSR method described earlier. In Gallup’s experiment (Gallup, 1970), chimpanzees performed mark-directed behavior by touching bright marks on their faces that were visible only in the mirror. A similar approach, but in a different modality, is implemented in the so-called “olfactory mirror” method aimed at identifying the ability of animals to distinguish their odor traces from other odors, including conspecifics. To date, dogs (Horowitz, 2017) and gray wolves (Gatti et al., 2020), snakes (Burghardt, et al., 2021) and geckos (Szabo, Ringler, 2023) have passed the test. In particular, R. Cazzolla Gatti notes “scent-rolling behavior” in wolves (Gatti et al., 2020).
When the behavior is oriented to other objects (Fig. 2, yellow-orange blocks), the subject’s own body (and consideration of its parameters) is not the goal, but the means. This manifestation of self-representation is studied using the “Body-as-obstacle” task. The experimental design presupposes that in order to solve an external task (for example, getting bait), the animal needs to consider the parameters of its body (size, weight, etc.), which creates an obstacle for implementation of the goal-directed behavior. The ability to consider one’s own weight was revealed in children aged 18-26 months (Brownell et al. 2007) and Indian elephants (Dale, Plotnik, 2017), domestic dogs (Lenkei et al., 2021) and rats (Khvatov, Kharitonov, 2021). The ability to take into account the size of their body was discovered in children aged 18–26 months (Brownell et al. 2007), budgerigars (Schiffner et al., 2014), domestic dogs (Lenkei et al., 2020), snakes (Khvatov et al., 2019), gray crows (Khvatov et al., 2021), domestic ferrets (Khvatov et al., 2023), snakes (Khvatov et al., 2019), toads (Khvatov & Zhelankin, 2018) and rats (Khvatov et al., 2024).
We believe that the representation of one’s own body when orienting individual behavior toward other objects is a basic type in the above-mentioned modality. This means that both in the phylogenetic and ontogenetic development, animals first acquire the ability to identify the parameters of their own body considering them as possible obstacles to the implementation of their behavior, and only under this condition that they can have motivation for purposeful self-study
and/or self-modification. Accordingly, from the point of view of reveiling the features of self-awareness, the body-as-obstacle method has greater power in comparison with MSR. This thesis may be confirmed by the fact that in the overwhelming majority of cases of using the body-as-obstacle task, positive results were obtained: animals took into account the parameters of their own body, even if not on the first try, but were able to learn (e.g. toads, see: Khvatov & Zhelankin, 2018; and snakes (Khvatov et al., 2019)). In addition, the MSR task is more complex than direct consideration of one’s own body parameters, since it involves instrumental activity. The subject must indirectly interact with the body using the mirror as a tool, and this involves additional cognitive mechanisms.
The colored blocks in Fig. 3 represent the modalities of own body representation studied to date in animals of different species. In addition to the above data, the ability of humans to recognize their own voices (Kaplan et al., 2008) and mirror-induced self-directed behavior in mice (Yokose et al., 2023) were established.
Figure 3. Modalities of body representation studied to date in animals of different species (for explanations see the text).
The selection of modalities of self-representation of one’s own appearance based on the olfactory modality or the representation of one’s own smell based on the auditory modality. For a further grounding of the model, we additionally introduce the category of “self-representation module”. The self-representation module is a subsystem, i.e. a relatively autonomous structural unit of representation of one’s own body, specialized in solving a number of similar behavioral problems and formed by means of integration of several different modalities. Accordingly, the modules of self-representation correspond to the properties identified by J. Fodor (1983), in particular, domain specificity, informational encapsulation, and fast speed of data processing. On the other hand, such a definition of the self-representation module is close to the understanding of “a functional organ” by A.A. Ukhtomsky (Ukhtomsky, 1927) and “a perceptual system” within the framework of the ecological approach (Gibson, 1979).
In turn, the evolutionary emergence of various self-representation modules occurs as a result of the mechanism of exaptation (Gould & Vrba, 1982) of the functions of individual sensory and effector systems of animals in accordance with the changing features of their ecology and behavior. Accordingly, different animal species may have different (both qualitatively and quantitatively) sets of self-representation modules, similarly to a Swiss army knife that contains many various tools (a metaphor proposed by Cosmides and Tooby, 1994). Further we consider several examples of such subsystems.
One of the most obvious illustrations will be humans. We have a highly integrated (in terms of its modal structure) visual-tactile-auditory module of representation of our own body, providing great behavioral flexibility and a high degree of abstraction of self-awareness (Fig. 4). This thesis is substantiated by the following. First, a person has a visual-tactile synthesis: visual and tactile systems may provide similar cognitive processes to sensory inputs that enable humans to rely merely on one modality in the absence of another to recognize surrounding objects (Tabrik et al., 2021). Similarly, it has been demonstrated that visual recognition of appearance itself and recognition of one’s own voice have similar neural support (Kaplan et al., 2008). Secondly, a high correlation has been found between the results of the MSR and body-as-obstacle task tests in humans in the second half of the second year of life (Barth et al., 2004; Brownell et al., 2007; Moore et al., 2007).
Figure 4. Modal organization of the visual-tactile-auditory module in humans (for explanation see the text).
The emergence of the visual-tactile-auditory module of self-representation could have been caused by the long-term evolution of the tool activity of apes and hominids (Toth & Schick, 2015). In addition, the evolution of the mind of representatives of the genus Homo was associated with the growth of cognitive fluidity, due to which previously isolated modules (like tools in a Swiss army knife (Cosmides & Tooby, 1994)) were connected to each other, forming larger highly integrative structures (Mithen, 1996; 2013; 2023). It is appropriate to note that the idea of cognitive fluidity widely circulated in recent years is consistent with the basic provisions of the Differentiation-Integration Approach (Chuprikova 2007, 2009). According to it, the development of any system begins with a homogeneous undifferentiated whole, then giving way to greater differentiation and autonomy of individual components, and later coming to a new integration.
Of course, this module of human self-representation is not the only one. In particular, the olfactory modality and the perception of one’s own body smell deserve special attention, even taking into account the evolutionary reduction of olfaction. Nevertheless, the functional load of the visual-tactile-auditory self-representation subsystem is extremely high, which probably makes it the leading one in terms of regulation of human behavior. In particular, this may explain certain anthropocentrism and the presentation of passing the MSR test as a sign of very high development of self-awareness (de Waal, 2019).
Figure 5. Modal organization of the visual-tactile-auditory module in humans (for explanation see the text).
Indeed, the research results suggest that animals have a larger number of more differentiated and functionally specialized modules for representing their own bodies. Considering rodents (mice and rats) as an example, we have the reason to describe two such modules (Fig. 5). First, the subsystem of tactile accounting of the size and weight of one’s own body when interacting with surrounding objects (Khvatov, Kharitonov, 2021; Krupa et al., 2001). Second, mice perform mirror-induced self-directed behavior, that is, they demonstrate signs of recognizing their own appearance, but only in the presence of tactile stimulation from marks applied to their bodies (Yokose et al., 2023).
It should be emphasized that due to the paucity of empirical material available to date, the above descriptions of self-representation modules are only plausible hypotheses that require further verification and clarification. Nevertheless, the proposed concept of the modular-modal organization of the representation of one’s own body, firstly, allows for a consistent interpretation of the existing data. In addition, it has an heuristic value in the sense that it provides grounds for putting forward and testing hypotheses about the specificity of the representation of one’s own body in previously unstudied animal species, taking into account the peculiarities of their ecology, physiology of sensory and effector systems, as well as behavior. For example, we can assume the presence of a module for representing various parameters of one’s own body (when focusing on external objects) based on the auditory modality in bats, taking into account their echolocation system (Müller, 2004) (Fig. 6).
Figure 6. Self-representation module of bats based on auditory modality (see text for explanations).
Based on the above provisions of the model of modal-modular representation of one’s own body, below we formulate a number of requirements for experimental procedures that allow us to identify the specificity of functioning of self-representation subsystems in individual representatives of the animal kingdom.
One should take into account that the data on the features of animal representation of their bodies accumulated to date are rather scarce, and therefore many studies are exploratory. Accordingly, as noted earlier, the description of self-representation modules in many animals is to a major extent hypothetical. Nevertheless, it is necessary to clearly define the modality of the sensory input of self-representation and the modality of the body parameters taken into account, which are planned to be studied in representatives of a particular species, since the experimental procedure and setup directly depend on this. This should be done based on the available data on the features of ecology, morphophysiology of sensory and effector systems, as well as the behavior of the selected animal species. Even if studies of a certain self-representation module in the selected animal species have been previously conducted, understanding its modalities is necessary to avoid both methodological inaccuracies and interpretation errors.
The third modality deserves special attention, i.e. motivational orientation of the behavior regulation of which involves self-representation. Since the representation of one’s own body in the orientation of individual behavior to other objects is a basic type, it should first be established whether the animal is capable of considering as an obstacle certain parameters of its own body received by it through a certain sensory input. Accordingly, for this purpose it is necessary to use the “Body-as-obstacle task” methodology. Only after identifying this ability in a specific animal species is it advisable to study whether it demonstrates signs of a direct motivational orientation toward studying or modifying its own body. In this context, it is appropriate to recall the fair criticism of the MSR method in that many animals do not show such an interest in their own appearance as humans.
The Body-as-obstacle task technique involves animals demonstrating signs of their own body representation during interaction with surrounding objects to solve an external experimental task in which the goal is not their own body, but, for example, reaching a bait. The procedure for studying the features of animal’s representation of its body should include two stages. At the first stage, the animal, having encountered a certain experimental task, forms a certain behavioral algorithm for solving it in a series of trials. In addition, the experimental procedure should allow identifying predictors of the animal’s choice of a certain behavior in the current situation. At the second stage, the experimental setup is changed in such a way that the animal discovers the impossibility of satisfying its need with the previously developed behavioral algorithm. To modify its own behavior, the animal will need to take into account the physical parameters of its own body. In this case, the experimental plan should reveal precisely the animal’s ability to systematically choose (develop) the necessary behavior taking into account the features of its body, excluding the trial and error method.
To illustrate one possible realization of the above-mentioned theoretical considerations and requirements for experimental procedures of studying the characteristics of the representation of own body by animals, we present a method developed and repeatedly tested by us.
The subjects used were grey crows (Khvatov et al., 2021), domestic ferrets (Khvatov et al., 2023), and brown rats (Khvatov et al., 2024). Despite the great diversity, representatives of all species in their natural habitats are faced with tasks involving the penetration of various openings of their own body or its parts.
Figure 7. The experimental setup: 1—launch box; 2—finish box with bait; a, b, c—holes in the partition (Khvatov et al., 2023).
The experimental setup (see fig. 7) is a rectangular box divided into four parts by three partitions. The central partition has three holes, into which special inserts are placed, allowing the size and shape of the hole to be modified. The partitions at the edges have one large hole equipped with opaque impenetrable inserts, thereby forming the starting and finishing sections. The outer walls of the setup and the floor are made of light opaque organic glass, the partitions and inserts are made of opaque black organic glass. The specific sizes of the setup parts vary according to the species of test animals.
Experimental procedure comprises preliminary food deprivation. Before the test, the animal is placed in the starting box and the bait is placed in the finishing box. The experimenter then lifts the insert, allowing the subject to choose a hole to penetrate, after which the animal moves to the finishing box, where the bait is located. As soon as the subject takes the bait, the finishing compartment is closed with a partition, and the trial is over.
Two experimental series are conducted with animals of each species.
In the experimental trials of the first series, all three holes have different sizes (small, medium, and large), but all are permeable to the animals’ bodies. A hole of each size is located alternately on the left, in the center, and on the right 12 times, so the experimental session consists of 36 trials. The objective of the series is to identify predictors of the choice of a particular hole for penetration by the subject, i.e. its position in the partition (left, right, or center), as well as its size.
The purpose of the second experiment is to determine whether the animals can choose one passable hole out of the three, although the other two are larger but impassable. The experimental series consists of 72 trials. Two types of trials are used: test and background, alternating in such a way that two background trials followed one test. This is necessary to exclude the effect of learning.
In each of the 24 test trials, there is only one passable hole of a smaller area, and two impassable holes of a larger area. Two types of passable holes are used, i.e. round and square, and two types of impassable holes, i.e. rectangles with a greater height and rectangles with a greater width.
In each of the 48 background trials, two passable holes of a larger area and one impassable hole of a smaller area are used. Like in the test trials, the impassable holes are round and square, the passable holes are high narrow and low wide rectangles. The sizes of the holes also vary according to the species of animals.
In each trial, two dependent variable indices are registered, i.e. the first approach to the hole and the first attempt to pass through the hole. An “approach” is considered to be a situation when the distance between the tip of the muzzle or beak and the hole was no more than 1 cm. An “attempt” is considered to be a situation when the tip of the animal’s muzzle (or beak) penetrates the hole.
The conclusion about the animal’s ability to take into account the limits of its own body (that the animal plans its behavior using the subsystem for taking into account the size of its body) is made in the case of a reliable predominance of the first approaches followed by passage through a single penetrable hole in 24 test trials.
The results of the experiments on various species showed that three of the six crows (Khvatov et al. 2021), six ferrets (Khvatov et al., 2023) of the six tested and as well as six rats out of the six that took part in the experiments were able to take into account the boundaries of their own body when planning their behavior.
It should be noted that representatives of a number of other species, in particular amphibians (Khvatov & Zhelankin, 2018) and reptiles (Khvatov et al., 2019) may ignore the size of their bodies when entering holes without preliminary trial, but were able to learn this in a number of experimental trials. These facts substantiate an additional question about the heritability or acquisition of modules for representing one’s body during ontogenesis by different animals.
The repeated successful use of the method described above, in our opinion, not only evidences to its methodological validity, but also the validity of the theoretical model of modal-modular representation of one’s own body, of which this method is a realization.
1. Conclusion
The theory we propose models the phenomenon of animal representation of one’s own body in the form of a set of relatively autonomous modules, the specific qualities of which are revealed by means of considering three modalities: the type of parameters of one’s own body taken into account, the type of sensory input and the motivational orientation of the activity during which self-representation occurs.
As has been shown, this approach has a number of advantages. Firstly, the evolution of self-representation is now presented not as a monolithic linear process, but as a multitude of independent lines of development of forms of self-representation in different animal species in accordance with the unique features of their phylogeny, ecology and morphology. Such a model allows one to consistently generalize the accumulating empirical data on the features of body representation manifested by various animals. Secondly, based on the provisions of the proposed theoretical model, it is possible to formulate a number of requirements for the methodology of organization of the experiments aimed at studying the features of body representation in animals. In particular, one of such methods was developed by the authors and proved its effectiveness.
Funding: The research is supported by Russian Science Foundation, grant # 24-28-01049 (I.A.K) and Russian Ministry for Science and Education, project # 0138-2024-0012 (A.N.K.).
Credit author statement:
The authors declare no conflict of interest.
- Allen, M. R. (2007). Mirror self-recognition in a gorilla (Gorilla gorilla gorilla) Master thesis. Florida International University. doi:10.25148/etd.fi13101588
- Anderson, J. R. (1984). Monkeys with mirrors: Some questions for primate psychology. International Journal of Primatology, 5(1), 81–98. doi:10.1007/bf02735149
- Artoni, P., Chierici, M. L., Arnone, F., Cigarini, C., De Bernardis, E., Galeazzi, G. M., Minneci, D. G., Scita, F., Turrini, G., De Bernardis, M., & Pingani, L. (2021). Body perception treatment, a possible way to treat body image disturbance in eating disorders: a case-control efficacy study. Eating and Weight Disorders, 26(2), 499–514. doi:10.1007/s40519-020-00875-x
- Asendorpf, J. B., Warkentin, & V., & Baudonniere, P. M. (1996). Self-awareness and Other-awareness II: Mirror Self-recognition, social contingency awareness, and synchronic imitation. Developmental Psychology, 32(2), 313—321. doi:10.1037/0012-1649.32.2.313
- Barabanshchikov, V. A. (2002). Perception and event. Petersburg: Aleteya. [Barabanshchikov V. A. Vospriyatie i sobytie. SPb.: Aletejya, 2002].
- Barsalou, L. W. (2008). Grounded cognition. Annual Review of Psychology, 59, 617—645. doi: 10.1146/annurev.psych.59.103006.093639.
- Barsalou, L. W. (2010). Grounded cognition: past, present, and future. Topics in Cognitive Science, 2(4), 716—724. doi: 10.1111/j.1756-8765.2010.01115.x.
- Barth, J., Povinelli, D. J., & Cant, J.G.H. (2004). Bodily origins of Self. In D. Beike, J. Lampinen, & D. Behrend (Eds.), The Self and Memory. Psychology Press.
- Berti, A. & Frassinetti, F. (2000). When Far Becomes Near: Remapping of Space by Tool Use. of Cognitive Neuroscience, 12(3), 415—420. doi:10.1162/089892900562237
- Beulah, A. (1972). Mirror self-image reactions before age two. Developmental Psychobiology, 5(4), 297–305. doi:10.1002/dev.420050403
- Brownell, C. A., Zerwas, S., & Ramani, G. B. (2007). “So big”: the development of body self-awareness in toddlers. Child Development, 1426–1440. doi:10.1111/j.1467-8624.2007.01075.x
- Buniyaadi, A., Taufique, S. K. T., & Kumar, V. (2020). Self‐recognition in corvids: Evidence from the mirror‐mark test in Indian house crows (Corvus splendens). Journal of Ornithology, 161, 341–350. https://doi.org/10.1007/s10336-019-01730-2
- Burghardt, G. M., Partin, A. M., Pepper, H. E., Steele, J. M., Liske, S. M., Stokes, A. E., Lathan, A. N., Springer, C. M., & Jenkins, M. S. (2021). Chemically mediated self-recognition in sibling juvenile common gartersnakes (Thamnophis sirtalis) reared on same or different diets: evidence for a chemical mirror? Behaviour, 158, 1169–1191. doi:10.1163/1568539X-bja10131
- Carlson, T., Alvarez, G., Wu Daw-an Verstraten, F.A.J. (2010) Rapid Assimilation of External Objects Into the Body Schema. Psychological Science, 21(7), 1000—1005. doi:10.1177/0956797610371962
- Chuprikova, N. I. (2007). Mental development: The principle of differentiation. St. Petersburg: St. Petersburg. [Chuprikova N.I. Umstvennoe razvitie: Princip differenciacii. SPb.: Piter, 2007]
- Chuprikova, N. I. (2009). The universal differentiation and integration law of development as the basis of an interdisciplinary paradigm theory of development. Theory of development: A differentiation and integration paradigm. Moscow: Languages of Slavic Cultures, 7—16. [Chuprikova N.I. Vseobshchij universal’nyj differencionno-integracionnyj zakon razvitiya kak osnova mezhdisciplinarnoj paradigmal’noj teorii razvitiya. // Teoriya razvitiya: Differencionno-integracionnaya paradigma. M.: Yazyki slavyanskih kul’tur, 2009. S. 7–16]
- Coren, S. (2001). How to speak dog: mastering the art of dog-human communication. Atria Books.
- Coren, S. (2004). How dogs think: Understanding the Canine mind. New York: Free Press.
- Cosmides, L. & Tooby, J. (1994). Beyond intuition and instinct blindness: toward an evolutionarily rigorous cognitive science. Cognition, 50(1-3), 41-77. doi: 10.1016/0010-0277(94)90020-5. PMID: 8039372.
- Costantini, M., Frassinetti, F., Maini, M., Ambrosini, E., Gallese, V., & Sinigaglia, C. (2014). When a laser pen becomes a stick: remapping of space by tool-use observation in hemispatial neglect. Experimental Brain Research, 232(10), 3233—3241.
- Couchman, J. J. (2011). Self-agency in rhesus monkeys. Biology Letters, 8(1), 39–41. doi:10.1098/rsbl.2011.0536
- Dale, R. & Plotnik, J. M. (2017). Elephants know when their bodies are obstacles to success in a novel transfer task. Scientific Reports, 7. doi:10.1038/srep46309.
- de Vignemont, F. (2010). Body schema and body image – pros and cons. Neuropsychologia, 48(3), 669–680.
- de Waal, F. B. M. (2019). Fish, mirrors, and a gradualist perspective on self-awareness. PLoS Biology, 17(2). doi:10.1371/journal.pbio.3000112
- Delfour, F. & Marten, K. (2001). Mirror image processing in three marine mammal species: killer whales (Orcinus orca), false killer whales (Pseudorca crassidens) and California sea lions (Zalophus californianus). Behavioural Processes, 53(3), 181–190. doi:10.1016/s0376-6357(01)00134-6
- Denes, G. (1999). Disorders of body awareness and body knowledge. In Denes G, Pizzamiglio L (eds.). Handbook of Clinical and Experimental Neuropsychology. Psychology Press. pp. 497–506. doi:10.4324/9781315791272-28
- Di Vita, A., Boccia, M., Palermo, L., & Guariglia, C. (2016). To move or not to move, that is the question! Body schema and non-action oriented body representations: An fMRI meta- analytic study. Neuroscience & Biobehavioral Reviews, 68, 37–46.
- Dijkerman, H. C. & de Haan, E. (2007). Somatosensory processes subserving perception and action. Behavioral and Brain Sciences, 30(02),
- Dyson, B. J., Dunn, A. K., & Alain, C. (2010). Ventral and dorsal streams as modality-independent phenomena. Cognitive Neuroscience 1(1),
- Fabri, K. E. (2004). Fundamentals of zoopsychology (3rd ed.). Moscow: UMK “Psychology”. [Fabri, K. E. (2004). Osnovy zoopsihologii (3-e izd.). M.: UMK “Psihologiya”].
- Fein, S. & Spencer, S. J. (1997). Prejudice as self-image maintenance: Affirming the self through derogating others. Journal of Personality and Social Psychology. 73(1), 31–44. doi:10.1037/0022-3514.73.1.31
- Ferris, J. (2012). Self-awareness with a simple brain. Scientific American Mind, 23(5), 28–29.
- Filippova, G. G. (2012). Zoopsychology and comparative psychology. Moscow: Akademiya. [Filippova, G. G. (2012). Zoopsihologiya i sravnitel’naya psihologiya. M.: Akademiya].
- Fodor, J. A. (1983). The Modularity of Mind: An Essay on Faculty of Psychology, MIT.
- Fox, M. (1982). Are most animals “mindless automatons”? A reply to Gordon G. Gallup. American Journal of Primatology, 3, 341–343.
- Gallagher, S. (1986). Body image and body schema: A conceptual clarification. Journal of Mind and Behaviour, 7(4), 541–554.
- Gallup, G. G. Jr. (1970). Chimpanzees: Self recognition. Science, 167(3914), 86–87. doi:10.1126/science.167.3914.86
- Gallup, G. G. Jr. & Anderson, J. R. (2020). Self-recognition in animals: Where do we stand 50 years later? Lessons from cleaner wrasse and other species. Psychology of Consciousness: Theory, Research, and Practice, 7, 46–58.
- Garbarinia, F., Fossataroa, C., Bertia, A., Gindria, P., Romanod, D., Piaa, L., Gattaf, F., Maravitad, A., & Neppi-Modona, M. (2015). When your arm becomes mine: Pathological embodiment of alien limbs using tools modulates own body representation. Neuropsychologia, 70, 402-413. doi:10.1016/j.neuropsychologia.2014.11.008
- Gatti, R. C., Velichevskaya, A., Gottesman, B., & Davis, K. (2021). Grey wolf may show signs of self-awareness with the sniff test of self-recognition. Ethology Ecology & Evolution, 33(4), 444-467. doi:10.1080/03949370.2020.1846628
- Gibson, J. J. (1966). The Senses Considered as Perceptual Systems. Allen and Unwin, London.
- Gibson, J. J. (1979). The Ecological Approach to Visual Perception. Boston: Houghton Mifflin.
- Giglia, G., Pia, L., Folegatti, A., Puma, P., Fierro, B., Cosentino, G., Berti, A., & Brighina, F. (2015). Far Space Remapping by Tool Use: A rTMS Study Over the Right Posterior Parietal Cortex. Brain Stimulation, 8(4), 795-800. doi: https://doi.org/10.1016/j.brs.2015.01.412
- Gore, J. S. & Cross, S. E. (2011). Defining and Measuring Self-Concept Change. Psychological Studies, 56(1), 135–141. doi:10.1007/s12646-011-0067-0
- Gould, S. J. & Vrba, E. S. (1982). Exaptation—a Missing Term in the Science of Form. Paleobiology, 8(1), 4-15.
- Gozli, D. G. & Brown, L. E. (2011). Agency and Control for the Integration of a Virtual Tool into the Peripersonal Space. Perception, 40(11), 1309—1319.
- Hall, D. A. (2003). Auditory pathways: Are ‘what’ and ‘where’ appropriate? Current Biology, 13(10), R406–R408.
- Hant, G. T. (2004). About the nature of consciousness. From a cognitive, phenomenological and transpersonal point of view. Moscow: Publishing House of AST. [Hant, G. T. (2004). O prirode soznaniya. S kognitivnoj, fenomenologicheskoj i transpersonal’noj tochek zreniya. M.: Izd-vo AST].
- Hauser, M., Miller, C., Liu, K., & Gupta, R. (2001). Cotton-top tamarins (Saguinus oedipus) fail to show mirror-guided self-exploration. American Journal of Primatology, 137, 131–137. doi:10.1002/1098-2345(200103)53:3<131::AID-AJP4>3.0.CO;2-X
- Head, H. & Holmes, G. (1911). Sensory disturbances from cerebral lesions. Brain, 34(2–3), doi:10.1093/brain/34.2-3.102
- Hewitt, J. P. (2009). The Social Construction of Self-Esteem. In Lopez, Shane J.; Snyder, C.R. (eds.). Oxford Handbook of Positive Psychology. Oxford University Press. pp. 217–224.
- Hill, H. M., Webber, K., Kemery, A., Garcia, M., & Kuczaj, S. A. (2015). Can sea lions’ (Zalophus californianus) use mirrors to locate an object? International Journal of Comparative Psychology, doi:10.46867/ijcp.2015.28.00.08
- Horowitz, A. (2017). Smelling themselves: Dogs investigate their own odours longer when modified in an “olfactory mirror” test. Behavioural Processes, 143, 17-24. doi:10.1016/j.beproc.2017.08.001.
- Hotta, T., Komiyama, S., & Kohda, M. (2018). A social cichlid fish failed to pass the mark test. Animal Cognition, 21(1), 127–136. doi:10.1007/s10071-017-1146-y
- James, W. (1911). Psychology. St. Petersburg. [Dzhejms U. Psihologiya. SPb., 1911]
- Johnson-Frey, S. (2004). The neural bases of complex tool use in humans. Trends in Cognitive Sciences, 8(2), 71—78. doi:10.1016/j.tics.2003.12.002
- Kaplan, J. T., Aziz-Zadeh, L., Uddin, L. Q., & Iacoboni, M. (2008). The self across the senses: an fMRI study of self-face and self-voice recognition. Social Cognitive and Affective Neuroscience, 3(3), 218–223. doi:10.1093/scan/nsn014
- Kaplan, L. A., Wiegner, T. , Newbold, J. D., Ostrom, P. H., & Gandhi, H. (2008). Untangling the complex issue of dissolved organic carbon uptake: a stable isotope approach. Freshwater Biology, 53, 855-864. doi:101111/j.1365-2427.2007.01941x
- Keizer, A., Smeets, M. A., Dijkerman, H. C., Uzunbajakau, S. A., van Elburg, A., & Postma, A. (2013). Too fat to fit through the door: first evidence for disturbed body-scaled action in anorexia nervosa during locomotion. PLOS ONE, 8(5). doi:10.1371/journal.pone.0064602
- Khvatov, I. (2011). The concept of the origin of the psyche of A. N.Leontiev at the present stage of the development of science. Psychological Research, 4(15). https://doi.org/10.54359/ps.v4i15 .873 [Khvatov, I. (2011). Koncepciya proiskhozhdeniya psihiki A.N.Leont’eva na sovremennom etape razvitiya nauki. Psihologicheskie issledovaniya, 4(15). https://doi.org/10.54359/ps.v4i15.873].
- Khvatov, I.A.; Ganza, P.N.; Kharitonov, A.N.; Samuleeva, M.V. Wistar Male Rats (Rattus norvegicus domestica) Are Aware of Their Dimensions. Animals 2024, 14, 3384. https://doi.org/10.3390/ani14233384
- Khvatov, I. A., Kharitonov, A. N., & Sokolov, A. Y. (2021). Brown rats may learn awareness of their body weight when interacting with environmental objects. Cognitive Research, Artificial Intelligence and Neuroinformatics, Proceedings of the 9th International Conference on Cognitive Sciences, Intercognsci-2020/ Velichkovsky B. M., Balaban P. M., Ushakov V. L. Cham, Switzerland: Springer, 2021. Р.1358.
- Khvatov, I. A., Smirnova, A. A., Samuleeva, M. V., Ershov, E. V., Buinitskaya, S. D., & Kharitonov, A. N. (2021). Hooded crows (Corvus cornix) may be aware of their own body size. Frontiers in Psychology, 12. doi:10.3389/fpsyg.2021.769397.
- Khvatov, I. A., Sokolov, A. Y., & Kharitonov, A. N. (2019) Snakes elaphe radiata may acquire awareness of their body limits when trying to hide in a shelter. Behavioral Sciences, (9)67. doi:10.3390/bs9070067.
- Khvatov, I. A., Sokolov, A. Y., & Kharitonov, A. N. (2023). Ferrets (Mustela furo) are aware of their dimensions. Animals (Basel), 13(3), doi:10.3390/ani13030444.
- Khvatov, I. A., Zhelankin, R. V. (2018). Features of learning in toads Bufo viridis during behavior that requires taking into account the limits of their own bodies. Experimental Psychology, 11(4), 5-16. [Eksperimental’naâ psihologiâ (Russia)] doi: 10.17759/exppsy.2018110401.
- Kohda, M., Bshary, R., Kubo, N., Awata, S., Sowersby, W., Kawasaka, K., Kobayashi, T., & Sogawa, S. (2023). Cleaner fish recognize self in a mirror via self-face recognition like humans. Proceedings of the National Academy of Sciences, U S A, 120(7). doi: 10.1073/pnas.2208420120.
- Kohda, M., Hotta, T., Takeyama, T., Awata, S., Tanaka, H., & Asai, J.-y. (2019). If a fish can pass the mark test, what are the implications for consciousness and self-awareness testing in animals? PLoS Biology, 17(2). doi:10.1371/journal.pbio.3000021
- Krachun, С., Lurz, R., Mahovetz, L., & Hopkins, W. D. (2019). Mirror self-recognition and its relation – ship to social cognition in chimpanzees. Animal Cognition, 22(6), 1171–1183. doi:10.1007/s10071- 019-01309-7.
- Kraft, F. L., Forštová, T., Utku Urhan, A., Exnerová, A., & Brodin, A. (2017). No evidence for self-recognition in a small passerine, the great tit (Parus major) judged from the mark/mirror test. Animal Cognition, 20(6), 1049–1057. doi:10.1007/s10071-017-1121-7
- Krupa, D. J., Matell, M. S., Brisben, A. J., & Oliveira, L. M., Nicolelis, M. A. (2001). Behavioral properties of the trigeminal somatosensory system in rats performing whisker-dependent tactile discriminations. Journal of Neuroscience, 21(15), 5752-5763.
- Lei, Yanyu. (2023). Sociality and self-awareness in animals. Frontiers in Psychology, doi:10.3389/fpsyg.2022.1065638.
- Lenkei, R., Faragó, T., Kovács, D., Zsilák, В., & Pongrácz, Р. (2020). That dog won’t fit: body size awareness in dogs. Animal Cognition, 23, 337–350. doi:10.1007/s10071-019-01337-3
- Lenkei, R., Faragó, T., Zsilák, B., & Pongrácz, P. (2021). Dogs (Canis familiaris) recognize their own body as a physical obstacle. Scientific Reports, 11,
- Leontiev, A. N. (1981). Problems of mental development (4th ed.). Moscow: Publishing House of the Moscow University. [Leontiev, A. N. (1981). Problemy razvitiya psihiki (4-e izd.). M.: Izd-vo Moskovskogo universiteta].
- Leontiev, A. N. (2024). Lectures on general psychology. Moscow: Publishing House “Smysl”. [Leont’ev A. N. Lekcii po obshchej psihologii. Moskva, Smysl, 2024]
- A. N. (1975). Activity. Conscience. Personality. Moscow: Publishing House “Politizdat”. [Leontiev. A.N. Deyatel’nost’. Soznanie. Lichnost. M.: Politizdat, 1975].
- Levin, K. (2000). Field theory in the social sciences. [Teoriya polya v social’nyh naukah / Per. E. Surpina. — SPb.: Rech’, 2000].
- Lyon, P. (2015) The cognitive cell: bacterial behavior reconsidered. Frontiers in Microbiology, doi: 10.3389/fmicb.2015.00264
- Lyon, P., Keijzer, F., Arendt D., & Levin, M. (2021). Reframing cognition: getting down to biological basics. Philosophical Transactions of the Royal Society. B376: 20190750. https://doi.org/10.1098/rstb.2019.0750
- Ma, X., Jin, Y., Luo, B., Zhang, G., Wei, R., & Liu, D. (2015). Giant pandas failed to show mirror self-recognition. Animal Cognition, 18(3), 713–721. doi:10.1007/s10071-015-0838-4
- Maravita, A. & Iriki, A. (2004). Tools for the body (schema). Trends in Cognitive Sciences, 8 (2), 79–86. doi:10.1016/j.tics.2003.12.008
- Marfenin, N. N. (2016). A decentralized organism on the example of colonial hydroids. Interdisciplinary scientific and applied Journal “Biosphere”, 8(3), 315-337. [Marfenin N.N. Decentralizovannyj organizm na primere kolonial’nyh gidroidov // Mezhdisciplinarnyj nauchnyj i prikladnoj zhurnal «Biosfera», 2016, t. 8, No 3, S. 315-337].
- Markus, H. (1977). Self-schemata and processing information about the self. Journal of Personality and Social Psychology, 35(2), 63–78. doi:10.1037/0022-3514.35.2.63
- Mather, J. A. & Kuba, M. J. (2013). The cephalopod specialties: complex nervous system, learning, and cognition. Canadian Journal of Zoology, 91(6), 431–449. doi:10.1139/cjz-2013-0009
- Milner, A. D. & Goodale, M. A. (1995). The visual brain in action. Oxford University Press. 2d ed. (2006), pp. 157–181.
- Milner, A. D. & Goodale, M. A. (2008). Two visual systems re-viewed. Neuropsychologia, 46(3), 774 – 777.
- Mithen, S. J. (1996). The Prehistory of the Mind: A Search for the Origins of Art, Religion, and Science. London: Thames and Hudson, 1996.
- Mithen, S. J. (2013). The Cathedral Model for the Evolution of Human Cognition. Evolution of Mind, Brain, and Culture, edited by Gary Hatfield and Holly Pittman, Philadelphia: University of Pennsylvania Press, pp. 217-234. https://doi.org/10.9783/9781934536605.217
- Mithen, S.J. (2023). The evolutionary foundations of Neolithic thought: The invention of words, cognitive fluidity and objects as cognitive anchors. In L.E. Bennison-Chapman (ed.) Bookkeeping Without Writing. Early Administrative Technologies in Context. PIHANS CXXXIV. Netherlands Institute for the Near East.
- Moeller, B., Zoppke, H., & Frings, C. (2016). What a car does to your perception: Distance evaluations differ from within and outside of a car. Psychonomic Bulletin & Review, 23(3), 781-788. doi: 10.3758/s13423-015-0954-9.
- Moore, C., Mealiea, J., Garon, N., & Povinelli, D. J. (2007). The Development of Body Self- Awareness. Infancy, 11(2), 157–174.
- Müller, R. (2004). A numerical study of the role of the tragus in the big brown bat”. The Journal of the Acoustical Society of America. 116 (6): 3701–3712
- Norman, D. A. (2013). The design of everyday things (Revised and expanded editions ed.). Cambridge, MA London: The MIT Press.
- Paillard, J. (1999). Body Schema and Body Image-a Double Dissociation in deafferented patients. In G. Gantchev, S. Mori, & J. Massion (Eds.), Motor control today and tomorrow. Sofia, Bulgaria: Academic Publishing House.
- Pitron, V. & de Vignemont, F. (2017). Beyond differences between the body schema and the body image: insights from body hallucinations. Consciousness and Cognition, 53, 115–121. doi:10.1016/j.concog.2017.06.006
- Plotnik, J. M., de Waal, F.B.M., & Reiss, D. (2006). Self-recognition in an Asian elephant. Proceedings of the National Academy of Sciences, 103(45). doi:10.1073/pnas.0608062103
- Povinelli, D. J., Rulf, A. B., Landau, K. R., & Bierschwale, D. T. (1993). Self-recognition in chimpanzees (Pan troglodytes): distribution, ontogeny, and patterns of emergence. Journal of Comparative Psychology, 107(4), 347–372. doi:10.1037/0735-7036.107.4.347
- Prior, H., Schwarz, A., & Güntürkün, O. (2008). Mirror-induced behavior in the magpie (Pica pica): Evidence of self-recognition. PLOS Biology, 6(8). doi:10.1371/journal.pbio.0060202
- Rajala, A. Z., Reininger, K. R., Lancaster, K. M., & Populin, L. C. (2010). Rhesus monkeys (Macaca mulatta) do recognize themselves in the mirror: Implications for the evolution of self-recognition. PLOS ONE, 5(9). doi:10.1371/journal.pone.0012865
- Reiss, D. & Marino, L. (2001). Mirror self-recognition in the bottlenose dolphin: A case of cognitive convergence. Proceedings of the National Academy of Sciences, 98(10), 5937–5942. doi:10.1073/pnas.101086398
- Ritchie, J. B. & Carlson, T. A. (2013). Tool Integration and Dynamic Touch. Psychological Science, 4(6), 1066-8. doi:10.1177/0956797612459768
- Robinson, T. (2023). Mirror self-recognition in ghost crab (Ocypode quadrata). Animal Cognition, 26(5), 1539–1549. doi:10.1007/s10071-023-01800-2. PMID 37314594
- Rochat, P. (2003). Five levels of self-awareness as they unfold early in life. Consciousness and Cognition 12(4), 717–731. doi:10.1016/s1053-8100(03)00081-3
- Rogers, K. R. (1994). A look at psychotherapy. The formation of man. Moscow: Publishing group “Progress”, “Univers”, 1994. [Rodzhers K. R. Vzglyad na psihoterapiyu. Stanovlenie cheloveka. — M.: Izdatel’skaya gruppa «Progress», «Univers», 1994]
- Roma, P., Silberberg, A., Huntsberry, M., Christensen, C., Ruggiero, A., & Suomi, S. (2007). Mark tests for mirror self-recognition in Capuchin monkeys (Cebus apella) trained to touch marks. American Journal of Primatology, 69(9), 989–1000. doi:10.1002/ajp.20404
- Romanski, L. M., Tian, B., Fritz, J., Mishkin, M., Goldman-Rakic, P. S., & Rauschecker, J. P. (1999). Dual streams of auditory afferents target multiple domains in theprimate prefrontal cortex. Nature Neuroscience 2(12), 1131–1136.
- Schilder, Paul.The image and appearance of the human body. (1935). London.
- Schiffner, I., Vo, H. D., Bhagavatula, P. S., & Srinivasan, M. V. (2014). Minding the gap: in-flight body awareness in birds. Frontiers in Zoology, 11, 64. doi:10.1186/s12983-014-0064-y
- Shaffer, V. A. & Renner, M. J. (2000). Black-and-White Colobus Monkeys (Colobus guereza) do not Show Mirror Self-Recognition. International Journal of Comparative Psychology, 13(3). doi:10.46867/C4H01J
- Shannon, C. E. (1948). A mathematical theory of communication. Bell System Technical Journal, 27(3), 379–423. doi:10.1002/j.1538-7305.1948.tb01338.x.
- Stolin, V. V. (1983). Self-awareness of personality.: Publishing House of the Moscow University. [Stolin, V. V. (1983). Samosoznanie lichnosti. M.: Izd-vo Moskovskogo universiteta].
- Suarez, S. D & Gallup, G. G. (1981). Self-recognition in chimpanzees and orangutans, but not gorillas. Journal of Human Evolution, 10(2), 175–188. doi:10.1016/s0047-2484(81)80016-4
- Szabo, B. & Ringler, E. (2023). Geckos differentiate self from other using both skin and faecal chemicals: evidence towards self-recognition? Animal Cognition, 26, 1-9. doi:10.1007/s10071-023-01751-8.
- Tabrik, S., Behroozi, M., Schlaffke, L., Heba, S., Lenz, M., Lissek, S., Güntürkün, O., Dinse, H.R., & Tegenthoff, M. (2021). Visual and tactile sensory systems share common features in object recognition. eNeuro, 8(5). doi: 10.1523/ENEURO.0101-21.2021. PMID: 34544756; PMCID: PMC8493885.
- The Cambridge Declaration on Consciousness (Archive) 7 July 2012. Written by Philip Low and edited by Jaak Panksepp, Diana Reiss, David Edelman, Bruno Van Swinderen, Philip Low and Christof Koch. University of Cambridge.
- The New York Declaration on Animal Consciousness New York University, 19 April 2024. URL: https://sites.google.com/nyu.edu/nydeclaration/declaration
- Thompson, E. (2010). Mind in life: Biology, phenomenology, and the sciences of mind. Harvard University Press.
- Toth, N. & Schick, K. (2015). Chapter 14 – Evolution of Tool Use. In Basics in Human Editor(s): Michael P. Muehlenbein, Academic Press, 2015, Pages 193-208, https://doi.org/10.1016/B978-0-12-802652-6.00014-1.
- Turner, J. & Oakes, P. (1986). The significance of the social identity concept for social psychology with reference to individualism, interactionism and social influence. British Journal of Social Psychology, 25(3), 237–252. doi:10.1111/j.2044-8309.1986.tb00732.x
- Ukhtomsky, A. A. (1927). Physiology of the motor apparatus. Moscow: “Practical Medicine”. [Uhtomskij A. A. (1927). Fiziologiya dvigatel’nogo apparata. M.: «Prakticheskaya medicina»].
- Varela, F. J., Thompson, E. T., & Rosch, E. (1991). The embodied mind: Cognitive science and human experience. Cambridge, MA: The MIT Press.
- Walraven, V., van Elsacker, L., & Verheyen, R. (1995). Reactions of a group of pygmy chimpanzees (Pan paniscus) to their mirror images: evidence of self-recognition. Primates, 36(1), 145–150. doi:10.1007/bf02381922
- Warren, J. E., Wise, R. J., & Warren, J. D. (2005). Sounds Do-Able: Auditory-motor transformations and the posterior temporal plane. Trends in Neurosciences, 28(12), 636–643.
- Warren, Jr. W. H. & Whang, S. (1987) Visual guidance of walking through apertures: body-scaled information for affordances. Journal of experimental psychology: human perception and performance, 13(3),
- Wilson, M. (2002). Six views of embodied cognition. Psychonomic Bulletin and Review, 9(4), 625–636. doi:10.3758/BF03196322
- Yokose, J., Marks, W.D., & Kitamura, T. (2023). Visuotactile integration facilitates mirror-induced self-directed behavior through activation of hippocampal neuronal ensembles in mice. Neuron, 112(2), 306-318. doi: 10.1016/j.neuron.2023.10.022. Epub 2023 Dec 5. PMID: 38056456.
- Zatorre, R. B., Marc A., & Pierre B. P. (2002). Where is ‘where’ in the human auditory cortex? Nature neuroscience, 5, 905-9.
- Zinchenko, V. P. (1999). The problem of external and internal formation of the image of oneself and the world as the realization of consciousness. The world of psychology, 1, 97 – 104. [Zinchenko V. P. Problema vneshnego i vnutrennego stanovleniya obraza sebya i mira kak realizaciya soznaniya. Mir psihologii, 1, 97 – 104]
- Zorina, Z. A., Poletaeva, I. I. (2003). Zoopsychology: elementary thinking of animals: A textbook for universities. M.: Aspect Press. [Zorina, Z. A., Poletaeva, I. I. (2003). Zoopsihologiya: elementarnoe myshlenie zhivotnyh: Uchebnoe posobie dlya vuzov. M.: Aspekt Press]
References