Biochemical Correlates of Individual Differences in Coping Intelligence
Abstract
Abstract
As a working definition we shall consider Coping Intelligence to be the ability to cope well with stressful situations. This research focuses on a systematic review of the biochemical correlates of Coping Intelligence. Research issue is, how does stress change biochemical and neural parameters, ensuring the progressive, stable or regressive development of a person? A total of 29 studies of Nature-associated journals from 2008 to 2022 (Median = 2017, SD = 3.44) were analyzed and results were organized into three sections such as neurochemistry of stress in animals, neurochemistry of stress in humans, and neurochemistry of resilience in stressful situations. Inclusion criteria were: (1) the biochemical changes in the body in response to stressful situations and stress-related diseases; (2) biochemical and neuronal aspects of stress resistance; (3) acute and chronic stress; (4) various biomaterials such as blood, microbiota and hair. Exclusion criteria were: (1) the biological level of Coping Intelligence is not described, (2) the main lines of human development in stressful situations are not demonstrated. Researchers examined a wide range of biological indicators of Coping Intelligence such as BDNF, pro-inflammatory cytokines, COMT, CDH1, HTR3, TNF-a, TRL9, ADRA2A, and OPRM1. The neurochemistry of acute stress in animals (mice, rodents, fish) correlates with elevated levels of catecholamines, monoamines, and pro-inflammatory cytokines. Early life stress of mice increases corticosterone in response to acute and chronic stress in adults. The neurochemistry of acute physical stress in humans seems to be similar to the neurochemistry of acute stress in animals. The chronic stress in humans is related to elevated level of central and peripheral inflammation that disrupts a person’s physical and mental health (hypertension, irritable bowel syndrome, diabetes mellitus, depression, anxiety disorder, etc.). Early life stress in humans contributes to stress-related diseases in adults. Resilience is described as a neurochemical system with such functional aspects as balance, plasticity, rhythm, emotion regulation, and accumulation of experience. These findings generally supported the different lines of subject’s development in the stressful situations across the studies reviewed. It should be pointed out that individual differences in Coping Intelligence are manifested not only at the biological, but also at the psychological and sociocultural levels.
Introduction
Stress is defined as a state of threat to the homodynamic balance due to a wide range of internal or external, real or perceived challenges or stimuli, called stressors (Agorastos & Chrousos, 2022). To maintain this optimal homodynamic state within a physiological range, organisms developed a very complex Stress Coping System (SCS)that serves the body’s self-regulation and adaptability by redirecting energy according to current needs. Inadequate, excessive or prolonged stress can overwhelm the body’s regulatory capacity and adaptive resources and cause inappropriate responses and a chronically altered homodynamic state associated with impaired mental and physical health and life expectancy. Neuroendocrine responses to stress depend on time of day, type of stressor, and its duration that leads to a vulnerable phenotype with impaired stress reactivity (i.e., a cumulatively elevated long-term risk of mental and physical morbidity). Consequences that are often observed in patients with chronic inflammatory diseases, such as fatigue, depressed mood, sleep disturbances, loss of appetite, muscle atrophy, cachexic obesity, bone loss and hypertension, may be the result of energy deprivation caused by an overactive immune system.
Anxiety is a manifestation of the normal response to stress. Cognitive programs move from complex associative operations to the rapid retrieval of unconscious emotional memories acquired during previous threatening situations. These programs are activated automatically to promote survival, i.e. to prevent distraction during stressful situations, while the ability to seek and experience pleasure is reduced, food consumption is reduced, and sexual activity and sleep are suspended. Monoamines, cytokines, glutamate, GABA, and other central mediators play a key role in the normal stress response (Gold, 2015, p. 32).
A key outcome of stress is the structural remodeling of neural architecture, which may be a sign of successful adaptation, while the persistence of these changes after the end of the stress indicates a lack of resilience. Excitatory amino acids and glucocorticoids play a key role in these processes, along with a growing list of extra- and intracellular mediators that includes endocannabinoids and brain-derived neurotrophic factor (BDNF) (McEwen at.al., 2015, p. 1353).
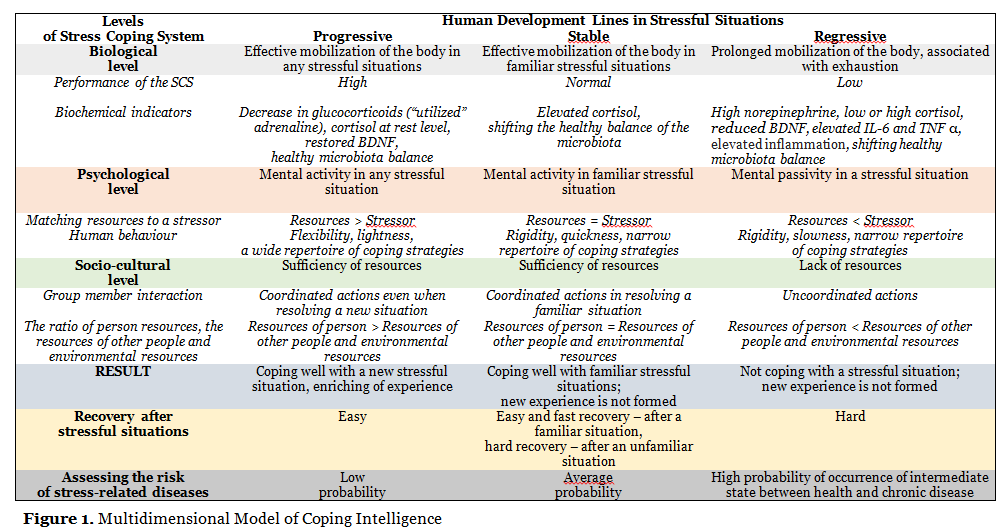
A review of the literature shows that the further the research advances in the neurochemistry of stress and the clearer becomes the understanding of the neurochemical basis of stress-resistant behavior, the more neurochemical systems associated with various stress-related diseases are found. There is a functional overlap of neurochemical systems responsible for the regulation of associated and not associated with the regulation of stress-resistant behavior. Neurochemical systems regulate each other, have multiple functions, and are represented by multiple neurotransmitters and mediators. Therefore, the question arises whether it is possible to unambiguously establish a correspondence between specific neurochemical systems and specific functional aspects of stress-resistant behavior, temperament traits, or symptoms of stress-associated diseases. We believe that this problem can be solved by studying the relationship between the neurochemical ensembles of the Stress-Coping System and the functional aspects of Coping Intelligence (Fig. 1).
It should be noted that despite the generally accepted thesis of the multidimensional nature of a person, modern studies of stress and coping persistently reproduce the classical patterns of reductionism in the knowledge of human behavior in difficult life situations. In a huge mass of fragmentary studies of various aspects of stress and coping, the idea of a holistic human nature was dissolved. Man as a microcosm embodies the entire multidimensionality of being, therefore, understanding the multidimensionality of human behavior in stressful situations should be associated with the analysis of many interacting systems, starting from the biochemical level and ending with the socio-cultural level within the framework of a holistic Stress Coping System.
Diverse epigenetic influences at an early age program each person for different trajectories of behavioral and physiological responses to later stressful life events (McEwen et al., 2015). Based on the Allostasis Model (Sterling, Eyer, 1988; Sterling, 2004), one can distinguish a progressive, stable, and regressive line of the Coping Intelligence development (Fig. 1).
The regressive development of the Coping Intelligence is manifested in the growth of the entropy of the Stress Coping System, in the reduction of its functionality, slowing down the speed of it functioning. There is a high probability of an intermediate state between health and chronic disease.
The stable functioning of the Coping Intelligence is realized in habitual situations that a person has previously encountered in his/her experience. The risk of occurrence of stress-related diseases is assessed as medium.
The progressive development of Coping Intelligence is due to the growth of integration of the Stress Coping System components, accompanied by a decrease in entropy. It manifests itself in the ease, speed and variability of coping, the emergence of original solutions to complex situations that were not previously presented in the individual experience of a person. The risk of stress-related diseases is low.
The congruence of Coping Intelligence systems provides an increase in organization (decrease in entropy) and an increase in human resilience, while incongruence leads to an increase in the entropy of the system and requires large expenditures of resources to achieve a useful result. In case of overcoming a stressful situation, a person acquires a new experience which reorganizes the Stress Coping System.
The biological level of Coping Intelligence reflects the biochemical and neuronal changes that occur during a stressful situation. The psychological level correlates with the individual psychological traits of a person, his/her cognitive assessment of a difficult life situation, the processes of conceptualization of resources, and the choice of coping strategies. The socio-cultural level covers the external context of human life, that is, a set of factors that determine the individual perception of stress and the choice of coping strategies (Kuvaeva, Volkova, 2022). When the actions of group members (family, organization, society, ethno-cultural group) are coordinated, a person overcomes a stressful situation; when the actions of group members are not coordinated, the resources of each individual within the collective subject are depleted up to its complete self-destruction (the death of civilization). The mediating factor between the biological and social levels of the Stress Coping System is the process of conceptualization of a stressful situation (the construction of the concept of Stress) and the regulation of behavior on its basis.
Method
Search and Screening
The search strategy included the use of the Nature e-journal database (nature.com). Nature is a weekly international journal publishing the finest peer-reviewed research in all fields of science and technology on the basis of its originality, importance, interdisciplinary interest, timeliness, accessibility, elegance and surprising conclusions. Nature also provides rapid, authoritative, insightful and arresting news and interpretation of topical and coming trends affecting science, scientists and the wider public. The first issue of Nature was published in November 1869. Two-year Impact Factor (2021) of Nature is 69.504.
Keywords that cover the neurochemical level of Coping Intelligence are presented in Table 1. Seven additional references were retrieved from the reference lists of the following papers in addition to the database search: Gold and Chrousos (Gold & Chrousos, 2002), Grayson (Grayson, 2016), Faraji (Faraji et al., 2017), Linz (Linz et al. 2019), Vandael and Gounko (Vandael & Gounko, 2019), Vindaz (Vindaz et al., 2018) and Zannas (Zannas et al., 2016).

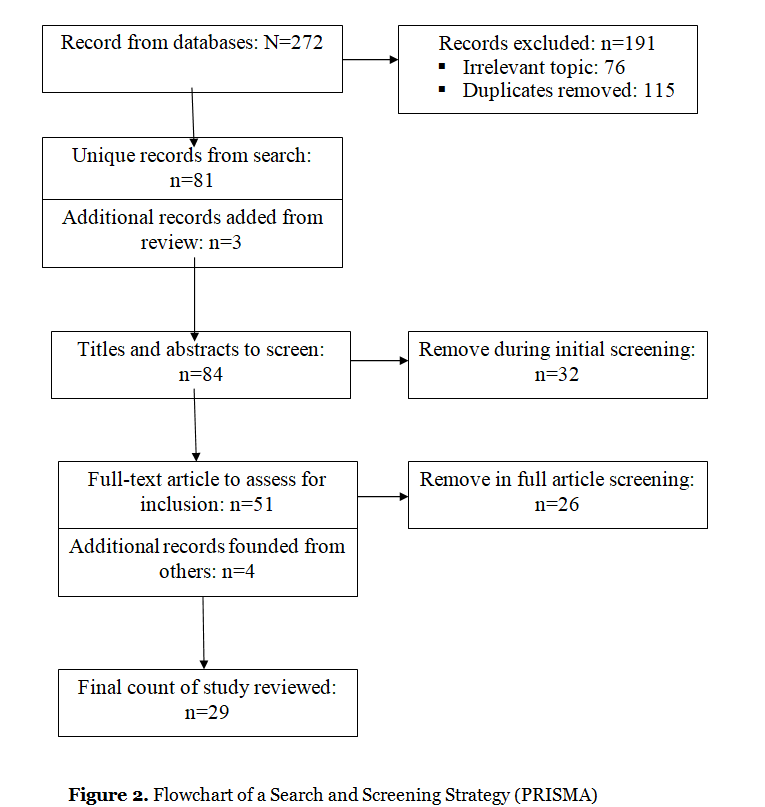
The four-phase PRISMA flow diagram (Figure 2) illustrates the search and screening process. Inclusion criteria were as follows. First, the sample included reviews that describe the biochemical changes in the body of animals and humans in response to stressful, difficult and intense situations. Second, some of the reviews are devoted to the biochemical and neuronal aspects of stress resistance, i.e. preservation of human health in the resolution of stressful situations. Another part of the reviews is devoted to the biochemical and neuronal aspects of stress-related diseases (rheumatic diseases, irritable bowel syndrome, mental disorders, oncological diseases, etc.). Third, the results described in the reviews were analyzed in terms of the type and duration of stress. The pool of reviewed reviews included studies examining the characteristics of biochemical changes in physical stressor, psychosocial stressor, and combined stressor (physical & social). Some research focuses on the biochemistry of acute stress. Reviews describing chronic stress often reflect the biochemistry of stress related diseases. Fourth, the study protocol included each of the following: (1) blood analysis, (2) microbiota analysis, (3) hair analysis, (4) description of the functioning of HPA axis and different brain regions. Fifth, the review articles are written in English, published over the past two decades (2002-2022) in Nature-associated journals. Therefore, reviews were excluded if they did not describe the biochemical level of Coping Intelligence and the main lines of human development in stressful situations (resilience vs stress-related diseases).
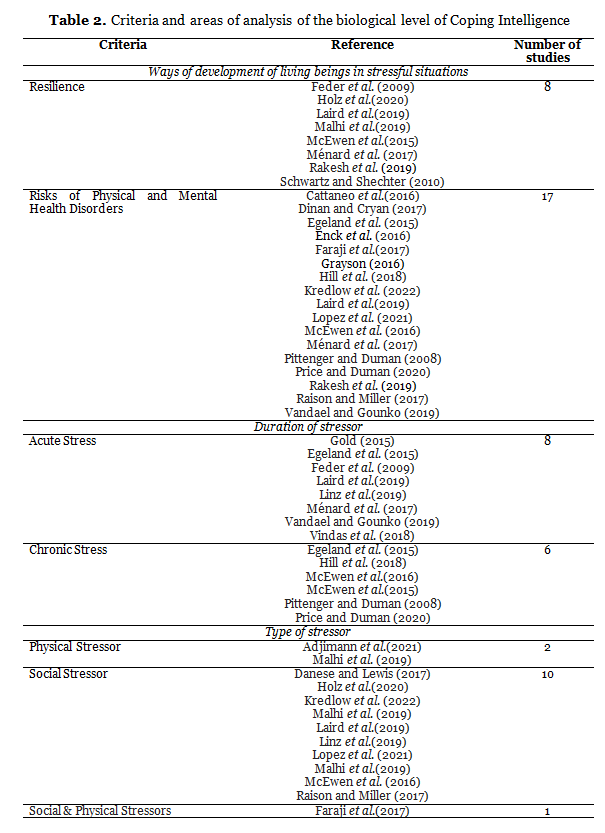
As shown in Table 2, our review included studies focusing primarily on the biochemistry of stress-related diseases (17 articles), while there were significantly fewer studies on stress resistance (8 articles). The review included an almost equal number of studies on acute (8) and chronic (6) stress, which allowed us to describe the neurochemistry of stress taking into account the duration of the stressor. Ten reviews describe data on the neurochemistry of stress induced by a psychosocial stressor; the neurochemistry of a physical stressor is considered less and this is done within the framework of an animal model.
Results
3.1. Neurochemistry of stress in animals
The literature describes in detail the neurochemical parameters of acute stress in rodents and mice. Acute stress manifests itself in elevated catecholamines (Kredlow et al., 2022), elevated corticosterone (Faraji et al, 2017; Feder et al., 2009), elevated serotonin (Feder et al., 2009), and elevated intracellular glutamate (McEwen et al., 2015). Similar results were obtained in a sample of juvenile gilthead seabream, which is characterized by an increase in the expression level of corticosterone, serotonin, dopamine, and BDNF under conditions of acute stress (Vindas et al., 2018).
Pro-inflammatory cytokines (Il-1B, Il-6) and TNF-α are released under acute stress in rodents (Ménard et al., 2017). Local and systemic increases in pro-inflammatory cytokines (Il-1, Il-6) in the microbiota cause activation of the hypothalamic-pituitary-adrenal axis (Dinan & Cryan, 2017). It was found that pro-inflammatory cytokines (Il-1beta), tumor necrosis factor (TNF-alpha), and interleukin Il-6 are modulated by stress (Rakesh et al., 2019). A study by Danese and Lewis showed that maternal separation stress in young primates and rats at an early age contributes to increased levels of pro-inflammatory cytokines (Il-1, Il-10) in adulthood (Danese & Lewis, 2017).
Changes in BDNF levels are indicators of chronic stress in rodents. It should be emphasized the inconsistency of data regarding brain-derived neurotrophic factor: Faraji (Faraji et al., 2017) found reduced BDNF in female rats under social isolation; Feder (Feder et al., 2009) found elevated levels of BDNF in animals.
According to the results of the study by McEwen, BDNF overexpression blocks the effects of chronic stress (McEwen et al., 2015).
Early life stress (ELS) in animals and multigenerational hereditary stress predetermine neurochemical changes in the body. If there was good maternal care, then the mice show low basal corticosterone levels and high corticosterone levels in response to the stressor. Rats that received poor maternal care showed elevated corticosterone in response to acute and chronic stressors. Glucocorticoids and HPA reactivity are also indicators of the quality of maternal care leading to better social and cognitive development. According to McEwen, “One measure of the efficacy of good maternal care is maternal stress self-regulation, referring to low basal CORT and a robust increased CORT secretion in response to a stressor” (McEwen et al., 2016, p. 9).
Under chronic stress, these “traumatized” mice show inflammation as well as an initial decrease in BDNF, which recovers to baseline levels after the stressor is overcome (McEwen et al., 2016). Multigenerational stress can reduce HPA axis activity to levels of stress resilience under repetitive stress (Faraji et al, 2017). COMT is an identified gene associated with traumatic stress response (Rakesh et al., 2019).
Features of the brain-gut interaction also determine the neurochemistry of stress and coping. The brain-gut-microbiota axis regulates immune responses and influences behavior. In the absence of stressors, the gut microbiota induces a steady state of ultra-low immune system activation; bacterial components stimulate macrophages and T-cells to produce pro-inflammatory cytokines such as Il-1β, tumor necrosis factor alpha (TNF-α) and Il-18. Elevated corticosterone in mice is reduced by feeding food containing Lactobacillus rhamnosus (JB-1) bacteria (Dinan & Cryan, 2017). Rats with reduced SERT expression and a prior history of maternal separation showed improved coping response to stress (Adjimann et al., 2021). It should be emphasized that social support reduces stress hormone levels in wild chimpanzees across stressful events and everyday affiliations (Wittig et al., 2016).
Under chronic stress conditions, rats with elevated peripheral Il-6 demonstrate learned helplessness (LH) behavior (Ménard et al., 2017) and passivity.
A developmental origin of individual differences in the responses to stress and implies that unknown early-life epigenetic influences program each individual to different trajectories of behavioral and physiological responses to later stressful life events (McEwen et al., 2016).
3.2. Neurochemistry of stress in humans
The latest research testifies a reciprocal regulation between adult neurogenesis and stress. That is, neurogenesis in adults can influence the response to stress, and stress can modulate levels of neurogenesis in adults. Neurogenesis affects whether new events are perceived as stressful or not. Stress, in turn, modulates the pathways of cytokines, neurotrophic factors, and signaling pathways of morphogens that determine neurogenesis. Undoubtedly, glucocorticoid hormones play the most important role in coping with stress (Egeland et al., 2015).
Acute stress in humans causes an increase in glucocorticoid levels (Egeland et al., 2015) and the release of pro-inflammatory cytokines (IL-1β, IL-6) and TRL-α (Ménard et al., 2017).
The psychosocial stressor in the form of performing the Triar Stress Test (TSST) causes an acute inflammatory response (Danese & Lewis, 2017). It was found in healthy participants (N=301, Mean Age = 40.65) during TSST that serum BDNF increases significantly after acute psychosocial stress and then decreases to baseline. Analysis of the dynamic interaction of BDNF and cortisol during performance of TSST showed an antagonistic relationship between BDNF and cortisol: higher peaks of BDNF after stress were associated with faster cortisol recovery; the magnitude of the cortisol response to stress was associated with faster recovery of BDNF after stress (Linz et al., 2019).
Chronic psychological stress is associated with inflammation and leukocytosis (Ménard et al., 2017), elevated levels of glucocorticoids (Pittenger & Duman, 2008), a change in BDNF expression as a function of stressor duration (Price & Duman, 2020); increased inflammation in the periphery and the brain (Il-1B, Il-6) (Egeland et al., 2015).
The literature emphasizes that chronic stress is a factor in the occurrence of various physical and mental diseases that have their own biochemistry. Cortisol as a biomarker for cardiovascular risk stratification correlates with obesity, hypertension, diabetes, metabolic syndrome, cardiovascular disease, and general anxiety disorder (Lopez et al., 2021). The high probability of progression of Alzheimer’s disease is correlated with hyperactivation of the HPA axis resulting from chronically high levels of stress (Vandael & Gounko, 2019). Post-traumatic stress disorder correlates with chronically elevated levels of catecholamines and corticotropin-releasing hormone (CRH), decreased cortisol, increased markers of inflammation (Hill et al., 2018). Elevated levels of systemic inflammation at an early age lead to a high risk of depression and psychosis at a young age (Danese & Lewis, 2017).
Depression is associated with higher levels of cortisol (Lopez et al., 2021), pro-inflammatory cytokines (Il-1β, Il-6), and TNFα (Ménard et al., 2017), decreased BDNF (Cattaneo et al., 2016; Price & Duman, 2020), maximal activation of the CRH/HPA axis and noradrenergic system (Gold, 2015). Analysis of the relationship between inflammation and depression revealed the expression of genes Il-1-α, Il-2, Il-3, Il-5, Il-8, Il-9, Il-10, Il-12A, Il-13, Il-15, and Il-18. People who are highly sensitive to social isolation have an increase in TRL-α in the blood and higher levels of pro-inflammatory cytokines (TNF и Il-6) (Raison & Miller, 2017). Patients with depression when perform TSST show significantly higher levels of Il-6 compared with the control group (Danese & Lewis, 2017). In depression, in addition to elevated levels of interleukin Il-6 and TNF α there is also an increase in the protein of the acute phase of C-reactive protein (Dinan & Cryan, 2017). An analysis of the regulatory cycles linking social stress and support revealed dopamine changes in such stress-related mental disorders as depression and PTSD. High cortisol response correlates with low pgACC activity as a regulator of negative emotions (Holz et al., 2020). Late-life depression correlates with decreased BDNF secretion (Laird et al., 2019).
Modern researchers consider irritable bowel syndrome as a multifactorial disease characterized by impaired immune function (Altered Immune Function: Il6, Il10, TNF); increased levels of serotonin in the blood plasma, increased level of BDNF, altered diversity and composition of the microbiota (Grayson, 2016). Irritable bowel syndrome is associated with such mental disorders as depression, anxiety, and neuroticism (Enck et al., 2016). It should be noted that disturbed microbiota is associated with various diseases (necrotizing enterocolitis in infants, obesity, diabetes, metabolic syndrome, irritable bowel syndrome, inflammatory bowel disease) (Dinan & Cryan, 2017).
Genetics, early life stress and living conditions determine neurochemical changes in the human body. The FKBP5 genetic biomarker is a modulator of glucocorticoid signaling and is associated with childhood abuse. Child physical abuse is associated with an increase in the HPA axis and inflammatory reactions; emotional abuse delays physiological recovery from stress and prolongs the effects of glucocorticoids (Malhi et al., 2019). Individuals with the FKBP5 “risk” allele are prone to developing post-traumatic stress disorder (Kredlow et al., 2022), major depressive disorder (MDD), and aggressive behavior (Zannas et al., 2016).
An analysis of hair cortisol concentrations as a summary and chronological indicator of children’s exposure to adverse experiences and other contextual stressors showed that higher cortisol in preschool children is associated with impaired socio-emotional development and an increased risk of developmental delay. Elevated cortisol in children aged 6-8 years is associated with a higher body mass index in girls and somatic complaints in boys. A higher concentration of cortisol in the hair was found in children with epilepsy and anorexia nervosa (Lopez et al., 2021). Increased cortisol and increased inflammation were observed in children who live in families with low socioeconomic status (Malhi et al., 2019).
Inndividual differences in neuroimmune and neuroendocrine shifts in response to prolonged stress likely shape behavioral vulnerability or resilience (Ménard et al., 2017).
3.3. Neurochemistry of resilience in stressful situations
Resilience is a dynamic multi-level system between neuroscience and behavior (Holz et. al., 2019). The latest data testified that up to two-thirds (65.7%) of people undergoing adversity remain relatively unscathed and demonstrate resilience in stressful situations (Malhi et. al., 2019). Resilience as an active process implies constant adaptive plasticity without external intervention (McEwen et al., 2015).
The resilience is understood as a multidimensional, dynamic capacity influenced by life-long interactions between internal and environmental resources (Laird et al., 2019; Malhi et. al., 2019). Internal resilience factors include plasticity, positive coping styles, optimism, and social support (Holz et. al., 2019). The resource of the environment is to create stressful situations and develop a positive adaptation of the body (Malhi et. al., 2019).
Animals that exhibit active behavior in response to environmental challenges have a lower glucocorticoid response compared to animals that exhibit passive behavior (Feder et. al., 2009). In humans, glucocorticoid and mineralocorticoid receptors are in an antagonistic relationship: low or high levels of cortisol cause an increase in mineralocorticoid receptors; glucocorticoid receptors reduce cell reproduction (Egeland et. al., 2015). The hormone oxytocin can reduce the activation of the HPA axis, causing an antidepressant effect (Menard et al., 2017).
Suppression of pro-inflammatory cytokines may be a marker of resilience to stress. Increased peripheral inflammation may be a predictor or consequence of stress tolerance, possibly because inflammatory markers influence behavioral outcomes through regulation of the HPA axis and hippocampal neurogenesis (Malhi et. al., 2019). Circulating immune cells maintenance hippocampal neurogenesis, spatial memory, brain-derived neurotrophic factor expression, and stress resilience (Schwartz & Shechter, 2010, p. 342).
Neuropeptide Y (NPY), as an endogenous mediator of resilience to stress-related mental illness, plays a key role in maintaining homeostasis and is involved in a variety of motivational, perceptual, and affective processes, including circadian rhythm, anxiety, appetite, alcohol use, and pain perception (Laird et al., 2019).
A review of the literature allows us to highlight such functional aspects of resilience as balance, plasticity, maintaining rhythms, relying on experience and its reassessment, and emotional regulation.
- Balance. Balance is manifested both in the interaction of different systems and within the system. For instance, resilience suggests a balance between neuropeptide (NPY) levels and CRH levels during stress. Resilience, understood as the ability to control stress-induced increases in CCR and cortisol through an elaborate negative feedback system, ensures optimal functioning and balance of glucocorticoid and mineralocorticoid receptors (Feder al., 2009). The interaction of the autonomic nervous system, the Hypothalamic-Pituitary-Adrenal (HPA) axis, and the immune system ensures dosing of the hormonal and inflammatory response to stress (Menard et al., 2017).
- Synaptic plasticity is associated with BDNF expression in the cortico-limbic regions of the brain (Malhi et. al., 2019). Increasing plasticity enables neuronal development, which in turn enhances adaptive capacity to resist challenge (Egeland et. al., 2015).
- Circadian rhythm disturbance is both a “stress factor” by increasing allostatic overload and a risk factor for other stressful experiences, highlighting the importance of timely action of glucocorticoids through the brain and body (Egeland et. al., 2015; McEwen et al., 2015).
Rhythmic HPA function seems to be necessary for normal initiation and termination of ACTH and other stress mediators. Disruption of normal oscillatory profiles of glucocorticoids could lead to desynchronized activity between different brain regions as well as peripheral organ systems. Epidemiologically, disrupted sleep and circadian rhythms lead to increased risk for development of psychiatric, cardiovascular or other physiological syndromes in shift workers. This dissonance is thought to contribute to several pathologies that are similar to the effects of chronic stress, including obesity and metabolic syndrome. Circadian (or diurnal) rhythms are a crucial factor that impact the stress response.
- The adaptive immune system can retain immunological memory of a stressor, thereby providing protection against future similar exposure to stress (Menard et al., 2017).
- Reappraisal of experience. Cognitive reappraisal of daily events leads to greater PFC and less amygdala activation to negative stimuli (Feder al., 2009).
- Emotion regulation. Emotional regulation is a more advanced fear-handling cognitive function, as amygdala is involved in the ability to process fearful stimuli, the hippocampus mediates the temporal and contextual aspects of fear. The basis of optimism such as the tendency to expect positive events in the future, is associated with stronger activation of amygdala and rostral ACC activation. The neural model of emotional regulation has ventral and dorsal systems (Holz et. al., 2019).
Researches of resilience at different ages are of great interest. In adolescence, there is a significant neuroplasticity of the brain, since it is at this age that the number of stressful situations increases significantly, individualization and dominance of relationships with peers occur. At the same time, major changes are taking place in systems that respond to stress (HPA axis). This means that as the social network expands, the sources of adversity become more common, while the resources needed to cope with current or potential future adversity remain limited (Malhi et. al., 2019). Analysis of the biopsychosocial correlates of resilience in late-life depression revealed that resilience correlates with biological factors such as the Val/Val allele, higher expression of mineralocorticoid receptors, higher BDNF methylation, higher levels of neuropeptide Y, efficient monoamine transfer, high levels of dehydroepiandrosterone (DHEA), moderate availability of estrogen. Resilience in old age is also correlated with psychosocial factors such as positive affectivity, active coping, social support, exercise and healthy eating, and secure attachment (Laird et al., 2019).
Thus, resilience is defined as an integrated process involving multiple peripheral and central mechanisms that promotes an adequate, non-pathological response to stress (Mehard et al., 2017). Two major factors, personality resilience and genetic differential susceptibility, determine the variability of an individual’s response to stress and developmental line: (1) inappropriate behavior and increased vulnerability to the onset of psychopathology, (2) development of resilience and adaptation (Malhi et. al., 2019).
Discussion
The presented review draws attention to the fact that biochemical markers of stress depend on the characteristics of the stressor and human resources. For example, cortisol is elevated major depressive disease and lower in general anxiety disorder, while in post-traumatic stress disorder, changes in cortisol depend on the type of traumatic experience and the time elapsed since the trauma (Lopez et al., 2021). There is evidence that BDNF expression varies greatly depending on the duration of the stressor (Egeland et al., 2015). Analysis of studies on the neurochemistry of stress in humans, as opposed to animal studies, suggests the impact of a psychosocial stressor on physical well-being and mental well-being. While in animal studies, the effect of physical threatening exposure is most often studied (the exception is the separation of mice and rats from their mother in early age). The accumulated stress biochemical effects contribute to the occurrence of stress-related diseases, including mental disorders (major depression, Alzheimer’s disease, etc.). These are diseases that cannot be studied in animals. Survey studies of biomarkers of acute and chronic stress make it possible to identify the neurochemical ensembles of Coping Intelligence and predict the risks of specific stress-related diseases.
Our analysis of reviews depicted to the neurochemistry of stress allowed us to put forward a hypothesis about sensitive periods in the development of Coping Intelligence. The first sensitive period falls on infancy and is associated with the functioning of the HPA axis. Early weaning from the mother contributes to the hyperactivation of the Stress Coping System; safe and emotionally comfortable infancy ensures the normal/adequate functioning of the Stress Coping System. As research shows that children raised in orphanages and having disorganized attachment in infancy have a large amygdala, which is a hypothetical biomarker for difficulty in emotional regulation of stressful situations (Laird et al., 2019). The second sensitive period in the development of Coping Intelligence is adolescence. Moderate stress in the life of a teenager contributes to the acquisition of adaptive responses to stress due to the increasing neuroplasticity of the brain (Malhi et. al., 2019).
In our opinion, there is probably another sensitive period in the development of Coping Intelligence, this is the period of early adulthood and middle age, when professional activity and family creation are associated with various difficulties, which can trigger prolonged inflammation in the body. This assumption requires additional research.
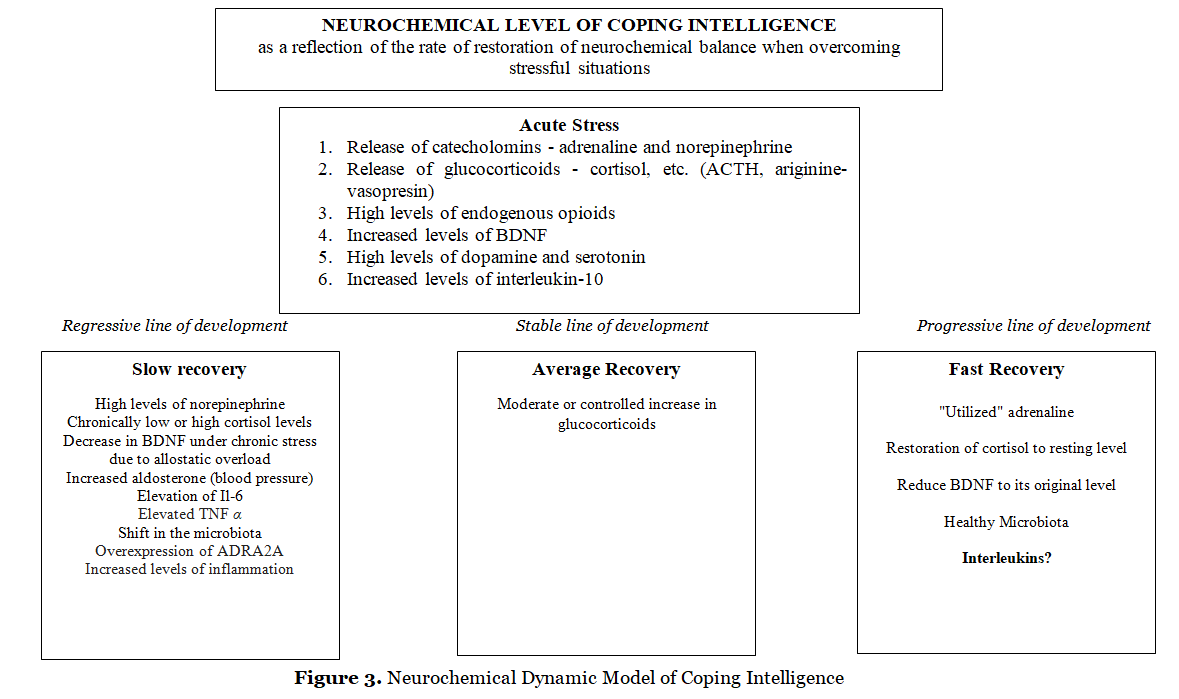
A possible neurochemical dynamic model of the Coping Intelligence is shown in fig. 3. A stressful situation leads to the activation of the sympathoadrenal system (SAS). The release of catecholamines (adrenaline, norepinephrine) provides a quick mobilization of the body and prepares it for action (“fight or flight”). Adrenaline initiates the release of insulin, which enhances glucose uptake by actively functioning organs. As a result of coping with stress, the concentration of adrenaline in the blood drops, as it is absorbed by the muscles (“utilized” adrenaline). However, if the body does not perform active actions and a high level of adrenaline remains in the blood, then the hypothalamic-pituitary-adrenal (HPA) system is triggered. The release of glucocorticoids (cortisol, etc.) maintains a high energy level in the body.
It should be noted that the studies do not take into account the “strength/weakness” of the nervous system and the severity of modal-specific activity. At the same time, the threshold of sensitivity in people with a “strong” and “weak” nervous system (highly active and low active in terms of V.M. Rusalov) differs (Ratanova, 2008). Under the influence of physical stress in respondents with a “strong” nervous system, the concentration of cortisol significantly decreases, while in respondents with a “weak” nervous system, it significantly increases. Respiratory rate normalization in respondents with a “strong” nervous system, compared with respondents with a “weak” nervous system, is achieved more slowly, they report the cessation of physical activity more quickly, but after additional stimulation they have more resources to continue it (Plachinta, 1978). The second (“cortisol”) period of the stress reaction is associated with a long-term mobilization of the body in response to the stressor, leading to suppression of the activity of the immune and reproductive systems, slowing down the development of the body. The suppression of regenerative processes continues during the sluggish stress response (Nikolaeva, 2019, p. 100). Termination of the stress response is ensured by the fact that activated glucocorticoid receptors reduce the activity of the HPA system with the help of negative biofeedback. Chronically low or high levels of glucocorticoids indicate suboptimal adaptation; a moderate or controlled increase in glucocorticoids is about physical and mental health (Nikolaeva, 2019, p. 103). The endogenous opioid regulatory system
(EOS) is aimed at producing a high level of endogenous opioids, thus inhibiting the development of a stress state (Bakhchina, 2022).
Acute stress state, in addition to the release of catecholamines, glucocorticoids (Vindaz et.al., 2018) and endogenous opioids, is associated with an increase in the level of Il-10, neutrotrophic factor (BDNF) (Linz, Puhlmann, Apostolakou et. al., 2019; Szuhany, Bugatti, Otto, 2015), high dopamine and serotonin levels (Aston-Jones, Chiang, Alexinsky, 1991; Kameneva et. al., 2022; Matsumoto & Hikosaka, 2009). Recovery of neurochemical balance after coping with acute stress occurs at low, medium, and high rates. The low rate of restoration of the neurochemical balance is associated with increased noadrenaline, a shift in the daily dynamics of glucocorticoid hormones (Bakhchina, 2022), decreased BDNF (Cattaneo et. al., 2016), increased Il-6 and TNF α (Dinan, 2009), and overexpression of ADR2A (Hohoff et. al., 2013), increased inflammation and a shift in the microbiota (Miller & Raison, 2016; Passos et. al., 2015; Straud, 2017), which determine the regressive line of human development in stressful situations. A moderate or controlled increase in glucocorticoids correlates (Nikolaeva, 2019) with an average rate of recovery of neurochemical parameters (stable line of development). The rapid restoration of neurochemical parameters ensures the progressive development of a person in stressful situations, due to the “reacted” adrenaline, restoration of cortisol to baseline, and a decrease in BDNF (Linz, Puhlmann, Apostolakou et. al., 2019). It should be noted the lack of empirical data on the state of the microbiota and the concentration of interleukins when a person quickly recovers from stressful situations. Based on the foregoing, we put forward a Neurochemical Dynamic Model of Coping Intelligence, which is presented in Fig. 3. The empirical verification of this model is the goal of our further research.
Conclusions
Highlights:
- The congruence of Stress Coping System levels ensures a high Coping Intelligence.
- The stress response is manifested in the HPA axis activation, elevation in pro-inflammatory cytokines and BDNF.
- Significant gaps in knowledge remain about neurochemistry of resilience.
- Resilience is associated with rhythms, cycles and synchronization of SCS.
- The effects of Early Life Stress can be offset by a healthy gut microbiota.
CRediT author statement: All authors contributed to the article and approved the submitted version.
References
Agorastos, A. & Chroustos, G.P. (2022). The neuroendocrinology of stress: the stress-related continuum of chronic disease development. Molecular Psychiatry, 27, 502-513. https://doi.org/10.1038/s41380-021-01224-9
Adjimann, T.S., Argañaraz, C.V. & Soiza-Reilly, M. (2021). Serotonin-Related Rodent Models of Early-Life Exposure Relevant for Neurodevelopmental Vulnerability to Psychiatric Disorders. Translational Psychiatry,11, 280. https://doi.org/10.1038/s41398-021-01388-6
Bahchina, A.V. (2022). Psychophysiology of stress. In: Psychophysiology: Ed. Yu.I. Aleksandrov, 5th edition. SPb.: Peter, 362-380. [Bahchina, A.V. (2022). Psihofiziologiya stressa. V: Psihofiziologiya: pod red. Aleksandrova Yu.I. 5-e izdanie. SPb.: Piter. 2022. 528 s.]
Brown, H. & Esterházy, D. (2021). Intestinal immune compartmentalization: implications of tissue specific determinants in health and disease. Mucosal Immunology,14, 1259–1270. https://doi.org/10.1038/s41385-021-00420-8
Cattaneo, A., Cattane, N., Begni, V. Pariante, C.M. & Riva M.A. (2016). The human BDNF gene: peripheral gene expression and protein levels as biomarkers for psychiatric disorders. Translational Psychiatry,6, 3-10. https://doi.org/10.1038/tp.2016.214
Danese, A. & Lewis, S.J. (2017). Psychoneuroimmunology of Early-Life Stress: The Hidden Wounds of Childhood Trauma? Neuropsychopharmacology,42, 99–114. https://doi.org/10.1038/npp.2016.198
Dinan, T.G. & Cryan, J.F. (2017). Microbes, Immunity, and Behavior: Psychoneuroimmunology Meets the Microbiome. Neuropsychopharmacology,42, 178–192. https://doi.org/10.1038/npp.2016.103
Egeland, M., Zunszain, P.A. & Pariante, C.M. (2015). Molecular mechanisms in the regulation of adult neurogenesis during stress. Nature Reviews Neuroscience,16, 189–200. https://doi.org/10.1038/nrn3855
Enck, P., Aziz, Q., Barbara, G., Farmer, A.D., Fukudo, S., Mayer, E.A., Niesler, B., Quigley, E.M.M., Rajilic-Stojanovic, M., Schemann, M., Schwille-Kiuntke, J., Simren, M., Zipfel, S. & Spiller, R.C. (2016). Irritable bowel syndrome. Nature Reviews Disease Primers,2, 16014. https://doi.org/10.1038/nrdp.2016.14
Faraji, J., Soltanpour, N., Lotfi, H., Moeeini, R., Moharreri, A-R., Roudaki, S., Hossenini, S.A., Olson, D.M., Abdollahi, A-A., Soltanpour, N., Mohajerani, M.H. & Metz, G.A.S. (2017). Lack of Social Support Raises Stress Vulnerability in Rats with a History of Ancestral Stress. Scientific Reports,7, 5277. https://doi.org/10.1038/s41598-017-05440-8
Feder, A., Nestler, E.J. & Charney, D.S. (2009). Psychobiology and molecular genetics of resilience. Nature Reviews Neuroscience, 10, 446-457. https://doi.org/10.1038/nrn2649
Gold, P. W. (2015). The organization of the stress system and its dysregulation in depressive illness. Molecular psychiatry, 20(1), 32-47.
Grayson, M. (2016). Irritable bowel syndrome. Nature, 533, 101. https://doi.org/10.1038/533S101a
Hill, M.N., Campolongo, P., Yehuda, R. & Patel, S. (2018). Integrating Endocannabinoid Signaling and Cannabinoids into the Biology and Treatment of Posttraumatic Stress Disorder. Neuropsychopharmacology, 43, 80–102 https://doi.org/10.1038/npp.2017.162
Hohoff, C., Gorji A., Kaiser, S., Willscher, E., Korsching, E., Ambree, O., Arolt, V., Lesch, K-P., Sachser. N., Deckert, J. & Lewejohann, L. (2013). Effect of Acute Stressor and Serotonin Transporter Genotype on Amygdala First Wave Transcriptome in Mice. PLoS ONE, 8(3), 1-13. https://doi.org/10.1371/journal.pone.0058880
Holz, N. E., Tost, H., & Meyer-Lindenberg, A. (2020). Resilience and the brain: a key role for regulatory circuits linked to social stress and support. Molecular psychiatry, 25(2), 379-396. https://doi.org/10.1038/s41380-019-0551-9
Kredlow, A.M., Fenster, R.J., Laurent, E.S., Ressler, K.J., & Phelps, E. (2022). Prefrontal Cortex, Amygdala, and Threat Processing: Implications for PTSD. Neuropsychopharmacoogy,47, 247–259. https://doi.org/10.1038/s41386-021-01155-7
Kuvaeva, I.O. & Volkova, E.V. (2022) The multidimensional nature of Coping Intelligence. In: History, modernity and prospects for the development of psychology in the system of the Russian Academy of Sciences: Proceedings of the International Anniversary Scientific Conference dedicated to the 50th anniversary of the Institute of Psychology of the Russian Academy of Sciences, November 16–18, 2022, Moscow / Ed. ed. D. V. Ushakov, A. L. Zhuravlev, A. V. Makhnach, N. E. Harlamenkova, A. V. Yurevich, I. I. Vetrova. – M.: Publishing house “Institute of Psychology of the Russian Academy of Sciences”. 500-502. [Kuvaeva, I.O. &Volkova, E.V. (2022). Mnogomernaya priroda sovladayushchego intellekta / Istoriya, sovremennost’ i perspektivy razvitiya psihologii v sisteme Rossijskoj Akademii nauk: Materialy Mezhdunarodnoj yubilejnoj nauchnoj konferencii, posvyashchennoj 50-letiyu sozdaniya Instituta psihologii RAN, 16–18 noyabrya 2022 g., Moskva / Otv. red. D. V. Ushakov, A. L. ZHuravlev, A. V. Mahnach, N. E. Harlamenkova, A. V. YUrevich, I. I. Vetrova. – M.: Izd-vo «Institut psihologii RAN», 500- 502.]
Laird, K.T., Krause, B., Funes, C. & Lavretsky H. (2019). Psychobiological factors of resilience and depression in late life. Translational Psychiatry,9, 88. https://doi.org/10.1038/s41398-019-0424-7
Linz, R., Puhlmann, L.M.C., Apostolakou, F., Mantzou, E., Papassotiriou, I., Chrousus, C.P., Engert, V. & Simger, T. (2019). Acute psychosocial stress increases serum BDNF levels: an antagonistic relation to cortisol but no group differences after mental training. Neuropsychopharmacology,44, 1797–1804 https://doi.org/10.1038/s41386-019-0391-y
Lopez, M., Ruiz, M.O., Rovnaghi, C.R., Tam, G.K-Y., Hiscoux, J., Gotlib, I.M., Barr, D.A., Carrion, V.G. & Anand, K.J.S. (2021). The social ecology of childhood and early life adversity. Pediatric Research, 89, 353–367. https://doi.org/10.1038/s41390-020-01264-x
Malhi, G.S., Das, P., Bell, E., Mattingly, G. & Mannie, Z. (2019). Modelling resilience in adolescence and adversity: a novel framework to inform research and practice. Translational Psychiatry, 9, 316. https://doi.org/10.1038/s41398-019-0651-y
McEwen, B.S., Bowles, N.P., Gray, J.D., Hill, M.N., Hunter, R.G., Karatsoreos, I.N. & Nasca, C. (2015). Mechanisms of stress in the brain. Nature Neuroscience, 18, 1353-63. https://doi.org/10.1038/nn.4086
McEwen, B., Nasca, C. & Gray, J. (2016). Stress Effects on Neuronal Structure: Hippocampus, Amygdala, and Prefrontal Cortex. Neuropsychopharmacology,41, 3–23. https://doi.org/10.1038/npp.2015.171
Ménard, C., Pfau, M.L., Hodes, G.E. & Russo, S.J. (2017). Immune and Neuroendocrine Mechanisms of Stress Vulnerability and Resilience. Neuropsychopharmacology,42, 62–80. https://doi.org/10.1038/npp.2016.90
Miller, A.H. & Raison, C.L. (2016). The role of inflammation in depression: from evolutionary imperative to modern treatment target. Nature Reviewers Immunology, 16, 22-34. doi: 10.1038/nri.2015.5.
Nikolaeva, E.I. (2019). Psychophysiology: Textbook for universities. third generation standard. SPb.: Peter. 704 p. [Nikolaeva E. (2019). Psihofiziologiya: Uchebnik dlya vuzov. Standart tret’ego pokoleniya. SPb.: Piter. 704 s.]
Passos, I.C., Vasconcelos-Moreno, M.P., Costa, L.G., Kunz, M., Brietzke, E., Quevedo, J., Salum, G., Magalhaes., P.V., Kapczinski, F. & Kauer-Sant’Anna M. (2015). Inflammatory markers in post-traumatic stress disorder: a systematic review, meta-analysis, and meta-regression. Lancet Psychiatry, 2, 1002–1012. doi: 1016/S2215-0366(15)00309-0
Pittenger, C. & Duman, R. (2008). Stress, Depression, and Neuroplasticity: A Convergence of Mechanisms. Neuropsychopharmacology, 33, 88–109. https://doi.org/10.1038/sj.npp.1301574
Plachinta, M. A. (1978). The strength of the nervous system and the nature of autonomic reactions in untrained people during physical activity of different intensity. Questions of Psychology, 4, 37-42. [Plachinta, M. A. (1978). Sila nervnoj sistemy i harakter vegetativnyh reakcij u netrenirovannyh lyudej pri fizicheskoj nagruzke raznoj intensivnosti. Voprosy psihologii, 4, 37-42.]
Price, R.B. & Duman, R. (2020). Neuroplasticity in cognitive and psychological mechanisms of depression: an integrative model. Molecular Psychiatry, 25(3), 530-543. doi: 10.1038/s41380-019-0615-x.
Raison, C. & Miller, A. (2017). Pathogen–Host Defense in the Evolution of Depression: Insights into Epidemiology, Genetics, Bioregional Differences and Female Preponderance. Neuropsychopharmacology, 42, 5–27. https://doi.org/10.1038/npp.2016.194
Rakesh, G., Morey, R.A., Zannas, A.S., Malik, Z., Marx, C.E., Clausen, A.N., Kritzer, M.D. & Szabo, ST. (2019). Resilience as a translational endpoint in the treatment of PTSD. Molecular Psychiatry, 24, 1268-1283. doi: 10.1038/s41380-019-0383-7.
Ratanova, T.A. (2008). Psychophysical scaling. Power of sensations, power of the nervous system and sensitivity. Moscow. Second edition. [Ratanova, T.A. (2008). Psihofizicheskoe shkalirovanie. sila oshchushchenij, sila nervnoj sistemy i chuvstvitel’nost’. Moscow. 2 izdanie].
Schwartz, M. & Shechter, R. (2010). Protective autoimmunity functions by intracranial immunosurveillance to support the mind: The missing link between health and disease. Molecular Psychiatry,15, 342–354. https://doi.org/10.1038/mp.2010.31
Sterling, P. (2004). Principles of allostasis: optimal design, predictive regulation, pathophysiology, and rational therapeutics. In: Allostasis, gomeostasis, and the costs of psychological adaptation. Ed. By J. Schulkin, 17 – 64.
Sterling, P. & Eyer, J. (1988). Allostasis: A New Paradigm to Explain Arousal Pathology. In: Handbook of life stress, cognition and health / Edited by S.Fisher and J. Reason. John Wiley & Sons, 629-649.
Straub, R. H. (2017). The brain and immune system prompt energy shortage in chronic inflammation and ageing. Nature Reviewers Rheumatology,13, 743–751. https://doi.org/10.1038/nrrheum.2017.172
Vandael, D. & Gounko, N.V. (2019). Corticotropin releasing factor-binding protein (CRF-BP) as a potential new therapeutic target in Alzheimer’s disease and stress disorders. Transational Psychiatry, 9, 272. https://doi.org/10.1038/s41398-019-0581-8
Vindas, M.A., Fokos, S., Pavlidis, M., Höglund, E., Dionysopoulou, S., Ebbesson, L.O.E., Papandroulakis, N., & Dermon, C.R. (2018). Early life stress induces long-term changes in limbic areas of a teleost fish: the role of catecholamine systems in stress coping. Scientific Reports,8, 5638. https://doi.org/10.1038/s41598-018-23950-x
Wittig, R.M., Crockford, C., Weltring, A., Langergraber, K.E., Deschner, T. & Zuberbuhler, K. (2016). Social support reduces stress hormone levels in wild chimpanzees across stressful events and everyday affiliations. Nature Communications, 7, Article number 13361, 1-8. https://doi.org/10.1038/ncomms13361
Zannas, A.S., Wiechmann, T., Gassen, N.C. & Binder, E.B. (2016). Gene–Stress–Epigenetic Regulation of FKBP5: Clinical and Translational Implications. Neuropsychopharmacology,41, 261–274. https://doi.org/10.1038/npp.2015.235
Comments (0)
As a working definition we shall consider Coping Intelligence to be the ability to cope well with stressful situations. This research focuses on a systematic review of the biochemical correlates of Coping Intelligence. Research issue is, how does stress change biochemical and neural parameters, ensuring the progressive, stable or regressive development of a person? A total of 29 studies of Nature-associated journals from 2008 to 2022 (Median = 2017, SD = 3.44) were analyzed and results were organized into three sections such as neurochemistry of stress in animals, neurochemistry of stress in humans, and neurochemistry of resilience in stressful situations. Inclusion criteria were: (1) the biochemical changes in the body in response to stressful situations and stress-related diseases; (2) biochemical and neuronal aspects of stress resistance; (3) acute and chronic stress; (4) various biomaterials such as blood, microbiota and hair. Exclusion criteria were: (1) the biological level of Coping Intelligence is not described, (2) the main lines of human development in stressful situations are not demonstrated. Researchers examined a wide range of biological indicators of Coping Intelligence such as BDNF, pro-inflammatory cytokines, COMT, CDH1, HTR3, TNF-a, TRL9, ADRA2A, and OPRM1. The neurochemistry of acute stress in animals (mice, rodents, fish) correlates with elevated levels of catecholamines, monoamines, and pro-inflammatory cytokines. Early life stress of mice increases corticosterone in response to acute and chronic stress in adults. The neurochemistry of acute physical stress in humans seems to be similar to the neurochemistry of acute stress in animals. The chronic stress in humans is related to elevated level of central and peripheral inflammation that disrupts a person’s physical and mental health (hypertension, irritable bowel syndrome, diabetes mellitus, depression, anxiety disorder, etc.). Early life stress in humans contributes to stress-related diseases in adults. Resilience is described as a neurochemical system with such functional aspects as balance, plasticity, rhythm, emotion regulation, and accumulation of experience. These findings generally supported the different lines of subject’s development in the stressful situations across the studies reviewed. It should be pointed out that individual differences in Coping Intelligence are manifested not only at the biological, but also at the psychological and sociocultural levels.
Stress is defined as a state of threat to the homodynamic balance due to a wide range of internal or external, real or perceived challenges or stimuli, called stressors (Agorastos & Chrousos, 2022). To maintain this optimal homodynamic state within a physiological range, organisms developed a very complex Stress Coping System (SCS)that serves the body’s self-regulation and adaptability by redirecting energy according to current needs. Inadequate, excessive or prolonged stress can overwhelm the body’s regulatory capacity and adaptive resources and cause inappropriate responses and a chronically altered homodynamic state associated with impaired mental and physical health and life expectancy. Neuroendocrine responses to stress depend on time of day, type of stressor, and its duration that leads to a vulnerable phenotype with impaired stress reactivity (i.e., a cumulatively elevated long-term risk of mental and physical morbidity). Consequences that are often observed in patients with chronic inflammatory diseases, such as fatigue, depressed mood, sleep disturbances, loss of appetite, muscle atrophy, cachexic obesity, bone loss and hypertension, may be the result of energy deprivation caused by an overactive immune system.
Anxiety is a manifestation of the normal response to stress. Cognitive programs move from complex associative operations to the rapid retrieval of unconscious emotional memories acquired during previous threatening situations. These programs are activated automatically to promote survival, i.e. to prevent distraction during stressful situations, while the ability to seek and experience pleasure is reduced, food consumption is reduced, and sexual activity and sleep are suspended. Monoamines, cytokines, glutamate, GABA, and other central mediators play a key role in the normal stress response (Gold, 2015, p. 32).
A key outcome of stress is the structural remodeling of neural architecture, which may be a sign of successful adaptation, while the persistence of these changes after the end of the stress indicates a lack of resilience. Excitatory amino acids and glucocorticoids play a key role in these processes, along with a growing list of extra- and intracellular mediators that includes endocannabinoids and brain-derived neurotrophic factor (BDNF) (McEwen at.al., 2015, p. 1353).
A review of the literature shows that the further the research advances in the neurochemistry of stress and the clearer becomes the understanding of the neurochemical basis of stress-resistant behavior, the more neurochemical systems associated with various stress-related diseases are found. There is a functional overlap of neurochemical systems responsible for the regulation of associated and not associated with the regulation of stress-resistant behavior. Neurochemical systems regulate each other, have multiple functions, and are represented by multiple neurotransmitters and mediators. Therefore, the question arises whether it is possible to unambiguously establish a correspondence between specific neurochemical systems and specific functional aspects of stress-resistant behavior, temperament traits, or symptoms of stress-associated diseases. We believe that this problem can be solved by studying the relationship between the neurochemical ensembles of the Stress-Coping System and the functional aspects of Coping Intelligence (Fig. 1).
It should be noted that despite the generally accepted thesis of the multidimensional nature of a person, modern studies of stress and coping persistently reproduce the classical patterns of reductionism in the knowledge of human behavior in difficult life situations. In a huge mass of fragmentary studies of various aspects of stress and coping, the idea of a holistic human nature was dissolved. Man as a microcosm embodies the entire multidimensionality of being, therefore, understanding the multidimensionality of human behavior in stressful situations should be associated with the analysis of many interacting systems, starting from the biochemical level and ending with the socio-cultural level within the framework of a holistic Stress Coping System.
Diverse epigenetic influences at an early age program each person for different trajectories of behavioral and physiological responses to later stressful life events (McEwen et al., 2015). Based on the Allostasis Model (Sterling, Eyer, 1988; Sterling, 2004), one can distinguish a progressive, stable, and regressive line of the Coping Intelligence development (Fig. 1).
The regressive development of the Coping Intelligence is manifested in the growth of the entropy of the Stress Coping System, in the reduction of its functionality, slowing down the speed of it functioning. There is a high probability of an intermediate state between health and chronic disease.
The stable functioning of the Coping Intelligence is realized in habitual situations that a person has previously encountered in his/her experience. The risk of occurrence of stress-related diseases is assessed as medium.
The progressive development of Coping Intelligence is due to the growth of integration of the Stress Coping System components, accompanied by a decrease in entropy. It manifests itself in the ease, speed and variability of coping, the emergence of original solutions to complex situations that were not previously presented in the individual experience of a person. The risk of stress-related diseases is low.
The congruence of Coping Intelligence systems provides an increase in organization (decrease in entropy) and an increase in human resilience, while incongruence leads to an increase in the entropy of the system and requires large expenditures of resources to achieve a useful result. In case of overcoming a stressful situation, a person acquires a new experience which reorganizes the Stress Coping System.
The biological level of Coping Intelligence reflects the biochemical and neuronal changes that occur during a stressful situation. The psychological level correlates with the individual psychological traits of a person, his/her cognitive assessment of a difficult life situation, the processes of conceptualization of resources, and the choice of coping strategies. The socio-cultural level covers the external context of human life, that is, a set of factors that determine the individual perception of stress and the choice of coping strategies (Kuvaeva, Volkova, 2022). When the actions of group members (family, organization, society, ethno-cultural group) are coordinated, a person overcomes a stressful situation; when the actions of group members are not coordinated, the resources of each individual within the collective subject are depleted up to its complete self-destruction (the death of civilization). The mediating factor between the biological and social levels of the Stress Coping System is the process of conceptualization of a stressful situation (the construction of the concept of Stress) and the regulation of behavior on its basis.
Search and Screening
The search strategy included the use of the Nature e-journal database (nature.com). Nature is a weekly international journal publishing the finest peer-reviewed research in all fields of science and technology on the basis of its originality, importance, interdisciplinary interest, timeliness, accessibility, elegance and surprising conclusions. Nature also provides rapid, authoritative, insightful and arresting news and interpretation of topical and coming trends affecting science, scientists and the wider public. The first issue of Nature was published in November 1869. Two-year Impact Factor (2021) of Nature is 69.504.
Keywords that cover the neurochemical level of Coping Intelligence are presented in Table 1. Seven additional references were retrieved from the reference lists of the following papers in addition to the database search: Gold and Chrousos (Gold & Chrousos, 2002), Grayson (Grayson, 2016), Faraji (Faraji et al., 2017), Linz (Linz et al. 2019), Vandael and Gounko (Vandael & Gounko, 2019), Vindaz (Vindaz et al., 2018) and Zannas (Zannas et al., 2016).
The four-phase PRISMA flow diagram (Figure 2) illustrates the search and screening process. Inclusion criteria were as follows. First, the sample included reviews that describe the biochemical changes in the body of animals and humans in response to stressful, difficult and intense situations. Second, some of the reviews are devoted to the biochemical and neuronal aspects of stress resistance, i.e. preservation of human health in the resolution of stressful situations. Another part of the reviews is devoted to the biochemical and neuronal aspects of stress-related diseases (rheumatic diseases, irritable bowel syndrome, mental disorders, oncological diseases, etc.). Third, the results described in the reviews were analyzed in terms of the type and duration of stress. The pool of reviewed reviews included studies examining the characteristics of biochemical changes in physical stressor, psychosocial stressor, and combined stressor (physical & social). Some research focuses on the biochemistry of acute stress. Reviews describing chronic stress often reflect the biochemistry of stress related diseases. Fourth, the study protocol included each of the following: (1) blood analysis, (2) microbiota analysis, (3) hair analysis, (4) description of the functioning of HPA axis and different brain regions. Fifth, the review articles are written in English, published over the past two decades (2002-2022) in Nature-associated journals. Therefore, reviews were excluded if they did not describe the biochemical level of Coping Intelligence and the main lines of human development in stressful situations (resilience vs stress-related diseases).
As shown in Table 2, our review included studies focusing primarily on the biochemistry of stress-related diseases (17 articles), while there were significantly fewer studies on stress resistance (8 articles). The review included an almost equal number of studies on acute (8) and chronic (6) stress, which allowed us to describe the neurochemistry of stress taking into account the duration of the stressor. Ten reviews describe data on the neurochemistry of stress induced by a psychosocial stressor; the neurochemistry of a physical stressor is considered less and this is done within the framework of an animal model.
3.1. Neurochemistry of stress in animals
The literature describes in detail the neurochemical parameters of acute stress in rodents and mice. Acute stress manifests itself in elevated catecholamines (Kredlow et al., 2022), elevated corticosterone (Faraji et al, 2017; Feder et al., 2009), elevated serotonin (Feder et al., 2009), and elevated intracellular glutamate (McEwen et al., 2015). Similar results were obtained in a sample of juvenile gilthead seabream, which is characterized by an increase in the expression level of corticosterone, serotonin, dopamine, and BDNF under conditions of acute stress (Vindas et al., 2018).
Pro-inflammatory cytokines (Il-1B, Il-6) and TNF-α are released under acute stress in rodents (Ménard et al., 2017). Local and systemic increases in pro-inflammatory cytokines (Il-1, Il-6) in the microbiota cause activation of the hypothalamic-pituitary-adrenal axis (Dinan & Cryan, 2017). It was found that pro-inflammatory cytokines (Il-1beta), tumor necrosis factor (TNF-alpha), and interleukin Il-6 are modulated by stress (Rakesh et al., 2019). A study by Danese and Lewis showed that maternal separation stress in young primates and rats at an early age contributes to increased levels of pro-inflammatory cytokines (Il-1, Il-10) in adulthood (Danese & Lewis, 2017).
Changes in BDNF levels are indicators of chronic stress in rodents. It should be emphasized the inconsistency of data regarding brain-derived neurotrophic factor: Faraji (Faraji et al., 2017) found reduced BDNF in female rats under social isolation; Feder (Feder et al., 2009) found elevated levels of BDNF in animals.
According to the results of the study by McEwen, BDNF overexpression blocks the effects of chronic stress (McEwen et al., 2015).
Early life stress (ELS) in animals and multigenerational hereditary stress predetermine neurochemical changes in the body. If there was good maternal care, then the mice show low basal corticosterone levels and high corticosterone levels in response to the stressor. Rats that received poor maternal care showed elevated corticosterone in response to acute and chronic stressors. Glucocorticoids and HPA reactivity are also indicators of the quality of maternal care leading to better social and cognitive development. According to McEwen, “One measure of the efficacy of good maternal care is maternal stress self-regulation, referring to low basal CORT and a robust increased CORT secretion in response to a stressor” (McEwen et al., 2016, p. 9).
Under chronic stress, these “traumatized” mice show inflammation as well as an initial decrease in BDNF, which recovers to baseline levels after the stressor is overcome (McEwen et al., 2016). Multigenerational stress can reduce HPA axis activity to levels of stress resilience under repetitive stress (Faraji et al, 2017). COMT is an identified gene associated with traumatic stress response (Rakesh et al., 2019).
Features of the brain-gut interaction also determine the neurochemistry of stress and coping. The brain-gut-microbiota axis regulates immune responses and influences behavior. In the absence of stressors, the gut microbiota induces a steady state of ultra-low immune system activation; bacterial components stimulate macrophages and T-cells to produce pro-inflammatory cytokines such as Il-1β, tumor necrosis factor alpha (TNF-α) and Il-18. Elevated corticosterone in mice is reduced by feeding food containing Lactobacillus rhamnosus (JB-1) bacteria (Dinan & Cryan, 2017). Rats with reduced SERT expression and a prior history of maternal separation showed improved coping response to stress (Adjimann et al., 2021). It should be emphasized that social support reduces stress hormone levels in wild chimpanzees across stressful events and everyday affiliations (Wittig et al., 2016).
Under chronic stress conditions, rats with elevated peripheral Il-6 demonstrate learned helplessness (LH) behavior (Ménard et al., 2017) and passivity.
A developmental origin of individual differences in the responses to stress and implies that unknown early-life epigenetic influences program each individual to different trajectories of behavioral and physiological responses to later stressful life events (McEwen et al., 2016).
3.2. Neurochemistry of stress in humans
The latest research testifies a reciprocal regulation between adult neurogenesis and stress. That is, neurogenesis in adults can influence the response to stress, and stress can modulate levels of neurogenesis in adults. Neurogenesis affects whether new events are perceived as stressful or not. Stress, in turn, modulates the pathways of cytokines, neurotrophic factors, and signaling pathways of morphogens that determine neurogenesis. Undoubtedly, glucocorticoid hormones play the most important role in coping with stress (Egeland et al., 2015).
Acute stress in humans causes an increase in glucocorticoid levels (Egeland et al., 2015) and the release of pro-inflammatory cytokines (IL-1β, IL-6) and TRL-α (Ménard et al., 2017).
The psychosocial stressor in the form of performing the Triar Stress Test (TSST) causes an acute inflammatory response (Danese & Lewis, 2017). It was found in healthy participants (N=301, Mean Age = 40.65) during TSST that serum BDNF increases significantly after acute psychosocial stress and then decreases to baseline. Analysis of the dynamic interaction of BDNF and cortisol during performance of TSST showed an antagonistic relationship between BDNF and cortisol: higher peaks of BDNF after stress were associated with faster cortisol recovery; the magnitude of the cortisol response to stress was associated with faster recovery of BDNF after stress (Linz et al., 2019).
Chronic psychological stress is associated with inflammation and leukocytosis (Ménard et al., 2017), elevated levels of glucocorticoids (Pittenger & Duman, 2008), a change in BDNF expression as a function of stressor duration (Price & Duman, 2020); increased inflammation in the periphery and the brain (Il-1B, Il-6) (Egeland et al., 2015).
The literature emphasizes that chronic stress is a factor in the occurrence of various physical and mental diseases that have their own biochemistry. Cortisol as a biomarker for cardiovascular risk stratification correlates with obesity, hypertension, diabetes, metabolic syndrome, cardiovascular disease, and general anxiety disorder (Lopez et al., 2021). The high probability of progression of Alzheimer’s disease is correlated with hyperactivation of the HPA axis resulting from chronically high levels of stress (Vandael & Gounko, 2019). Post-traumatic stress disorder correlates with chronically elevated levels of catecholamines and corticotropin-releasing hormone (CRH), decreased cortisol, increased markers of inflammation (Hill et al., 2018). Elevated levels of systemic inflammation at an early age lead to a high risk of depression and psychosis at a young age (Danese & Lewis, 2017).
Depression is associated with higher levels of cortisol (Lopez et al., 2021), pro-inflammatory cytokines (Il-1β, Il-6), and TNFα (Ménard et al., 2017), decreased BDNF (Cattaneo et al., 2016; Price & Duman, 2020), maximal activation of the CRH/HPA axis and noradrenergic system (Gold, 2015). Analysis of the relationship between inflammation and depression revealed the expression of genes Il-1-α, Il-2, Il-3, Il-5, Il-8, Il-9, Il-10, Il-12A, Il-13, Il-15, and Il-18. People who are highly sensitive to social isolation have an increase in TRL-α in the blood and higher levels of pro-inflammatory cytokines (TNF и Il-6) (Raison & Miller, 2017). Patients with depression when perform TSST show significantly higher levels of Il-6 compared with the control group (Danese & Lewis, 2017). In depression, in addition to elevated levels of interleukin Il-6 and TNF α there is also an increase in the protein of the acute phase of C-reactive protein (Dinan & Cryan, 2017). An analysis of the regulatory cycles linking social stress and support revealed dopamine changes in such stress-related mental disorders as depression and PTSD. High cortisol response correlates with low pgACC activity as a regulator of negative emotions (Holz et al., 2020). Late-life depression correlates with decreased BDNF secretion (Laird et al., 2019).
Modern researchers consider irritable bowel syndrome as a multifactorial disease characterized by impaired immune function (Altered Immune Function: Il6, Il10, TNF); increased levels of serotonin in the blood plasma, increased level of BDNF, altered diversity and composition of the microbiota (Grayson, 2016). Irritable bowel syndrome is associated with such mental disorders as depression, anxiety, and neuroticism (Enck et al., 2016). It should be noted that disturbed microbiota is associated with various diseases (necrotizing enterocolitis in infants, obesity, diabetes, metabolic syndrome, irritable bowel syndrome, inflammatory bowel disease) (Dinan & Cryan, 2017).
Genetics, early life stress and living conditions determine neurochemical changes in the human body. The FKBP5 genetic biomarker is a modulator of glucocorticoid signaling and is associated with childhood abuse. Child physical abuse is associated with an increase in the HPA axis and inflammatory reactions; emotional abuse delays physiological recovery from stress and prolongs the effects of glucocorticoids (Malhi et al., 2019). Individuals with the FKBP5 “risk” allele are prone to developing post-traumatic stress disorder (Kredlow et al., 2022), major depressive disorder (MDD), and aggressive behavior (Zannas et al., 2016).
An analysis of hair cortisol concentrations as a summary and chronological indicator of children’s exposure to adverse experiences and other contextual stressors showed that higher cortisol in preschool children is associated with impaired socio-emotional development and an increased risk of developmental delay. Elevated cortisol in children aged 6-8 years is associated with a higher body mass index in girls and somatic complaints in boys. A higher concentration of cortisol in the hair was found in children with epilepsy and anorexia nervosa (Lopez et al., 2021). Increased cortisol and increased inflammation were observed in children who live in families with low socioeconomic status (Malhi et al., 2019).
Inndividual differences in neuroimmune and neuroendocrine shifts in response to prolonged stress likely shape behavioral vulnerability or resilience (Ménard et al., 2017).
3.3. Neurochemistry of resilience in stressful situations
Resilience is a dynamic multi-level system between neuroscience and behavior (Holz et. al., 2019). The latest data testified that up to two-thirds (65.7%) of people undergoing adversity remain relatively unscathed and demonstrate resilience in stressful situations (Malhi et. al., 2019). Resilience as an active process implies constant adaptive plasticity without external intervention (McEwen et al., 2015).
The resilience is understood as a multidimensional, dynamic capacity influenced by life-long interactions between internal and environmental resources (Laird et al., 2019; Malhi et. al., 2019). Internal resilience factors include plasticity, positive coping styles, optimism, and social support (Holz et. al., 2019). The resource of the environment is to create stressful situations and develop a positive adaptation of the body (Malhi et. al., 2019).
Animals that exhibit active behavior in response to environmental challenges have a lower glucocorticoid response compared to animals that exhibit passive behavior (Feder et. al., 2009). In humans, glucocorticoid and mineralocorticoid receptors are in an antagonistic relationship: low or high levels of cortisol cause an increase in mineralocorticoid receptors; glucocorticoid receptors reduce cell reproduction (Egeland et. al., 2015). The hormone oxytocin can reduce the activation of the HPA axis, causing an antidepressant effect (Menard et al., 2017).
Suppression of pro-inflammatory cytokines may be a marker of resilience to stress. Increased peripheral inflammation may be a predictor or consequence of stress tolerance, possibly because inflammatory markers influence behavioral outcomes through regulation of the HPA axis and hippocampal neurogenesis (Malhi et. al., 2019). Circulating immune cells maintenance hippocampal neurogenesis, spatial memory, brain-derived neurotrophic factor expression, and stress resilience (Schwartz & Shechter, 2010, p. 342).
Neuropeptide Y (NPY), as an endogenous mediator of resilience to stress-related mental illness, plays a key role in maintaining homeostasis and is involved in a variety of motivational, perceptual, and affective processes, including circadian rhythm, anxiety, appetite, alcohol use, and pain perception (Laird et al., 2019).
A review of the literature allows us to highlight such functional aspects of resilience as balance, plasticity, maintaining rhythms, relying on experience and its reassessment, and emotional regulation.
- Balance. Balance is manifested both in the interaction of different systems and within the system. For instance, resilience suggests a balance between neuropeptide (NPY) levels and CRH levels during stress. Resilience, understood as the ability to control stress-induced increases in CCR and cortisol through an elaborate negative feedback system, ensures optimal functioning and balance of glucocorticoid and mineralocorticoid receptors (Feder al., 2009). The interaction of the autonomic nervous system, the Hypothalamic-Pituitary-Adrenal (HPA) axis, and the immune system ensures dosing of the hormonal and inflammatory response to stress (Menard et al., 2017).
- Synaptic plasticity is associated with BDNF expression in the cortico-limbic regions of the brain (Malhi et. al., 2019). Increasing plasticity enables neuronal development, which in turn enhances adaptive capacity to resist challenge (Egeland et. al., 2015).
- Circadian rhythm disturbance is both a “stress factor” by increasing allostatic overload and a risk factor for other stressful experiences, highlighting the importance of timely action of glucocorticoids through the brain and body (Egeland et. al., 2015; McEwen et al., 2015).
Rhythmic HPA function seems to be necessary for normal initiation and termination of ACTH and other stress mediators. Disruption of normal oscillatory profiles of glucocorticoids could lead to desynchronized activity between different brain regions as well as peripheral organ systems. Epidemiologically, disrupted sleep and circadian rhythms lead to increased risk for development of psychiatric, cardiovascular or other physiological syndromes in shift workers. This dissonance is thought to contribute to several pathologies that are similar to the effects of chronic stress, including obesity and metabolic syndrome. Circadian (or diurnal) rhythms are a crucial factor that impact the stress response.
- The adaptive immune system can retain immunological memory of a stressor, thereby providing protection against future similar exposure to stress (Menard et al., 2017).
- Reappraisal of experience. Cognitive reappraisal of daily events leads to greater PFC and less amygdala activation to negative stimuli (Feder al., 2009).
- Emotion regulation. Emotional regulation is a more advanced fear-handling cognitive function, as amygdala is involved in the ability to process fearful stimuli, the hippocampus mediates the temporal and contextual aspects of fear. The basis of optimism such as the tendency to expect positive events in the future, is associated with stronger activation of amygdala and rostral ACC activation. The neural model of emotional regulation has ventral and dorsal systems (Holz et. al., 2019).
Researches of resilience at different ages are of great interest. In adolescence, there is a significant neuroplasticity of the brain, since it is at this age that the number of stressful situations increases significantly, individualization and dominance of relationships with peers occur. At the same time, major changes are taking place in systems that respond to stress (HPA axis). This means that as the social network expands, the sources of adversity become more common, while the resources needed to cope with current or potential future adversity remain limited (Malhi et. al., 2019). Analysis of the biopsychosocial correlates of resilience in late-life depression revealed that resilience correlates with biological factors such as the Val/Val allele, higher expression of mineralocorticoid receptors, higher BDNF methylation, higher levels of neuropeptide Y, efficient monoamine transfer, high levels of dehydroepiandrosterone (DHEA), moderate availability of estrogen. Resilience in old age is also correlated with psychosocial factors such as positive affectivity, active coping, social support, exercise and healthy eating, and secure attachment (Laird et al., 2019).
Thus, resilience is defined as an integrated process involving multiple peripheral and central mechanisms that promotes an adequate, non-pathological response to stress (Mehard et al., 2017). Two major factors, personality resilience and genetic differential susceptibility, determine the variability of an individual’s response to stress and developmental line: (1) inappropriate behavior and increased vulnerability to the onset of psychopathology, (2) development of resilience and adaptation (Malhi et. al., 2019).
The presented review draws attention to the fact that biochemical markers of stress depend on the characteristics of the stressor and human resources. For example, cortisol is elevated major depressive disease and lower in general anxiety disorder, while in post-traumatic stress disorder, changes in cortisol depend on the type of traumatic experience and the time elapsed since the trauma (Lopez et al., 2021). There is evidence that BDNF expression varies greatly depending on the duration of the stressor (Egeland et al., 2015). Analysis of studies on the neurochemistry of stress in humans, as opposed to animal studies, suggests the impact of a psychosocial stressor on physical well-being and mental well-being. While in animal studies, the effect of physical threatening exposure is most often studied (the exception is the separation of mice and rats from their mother in early age). The accumulated stress biochemical effects contribute to the occurrence of stress-related diseases, including mental disorders (major depression, Alzheimer’s disease, etc.). These are diseases that cannot be studied in animals. Survey studies of biomarkers of acute and chronic stress make it possible to identify the neurochemical ensembles of Coping Intelligence and predict the risks of specific stress-related diseases.
Our analysis of reviews depicted to the neurochemistry of stress allowed us to put forward a hypothesis about sensitive periods in the development of Coping Intelligence. The first sensitive period falls on infancy and is associated with the functioning of the HPA axis. Early weaning from the mother contributes to the hyperactivation of the Stress Coping System; safe and emotionally comfortable infancy ensures the normal/adequate functioning of the Stress Coping System. As research shows that children raised in orphanages and having disorganized attachment in infancy have a large amygdala, which is a hypothetical biomarker for difficulty in emotional regulation of stressful situations (Laird et al., 2019). The second sensitive period in the development of Coping Intelligence is adolescence. Moderate stress in the life of a teenager contributes to the acquisition of adaptive responses to stress due to the increasing neuroplasticity of the brain (Malhi et. al., 2019).
In our opinion, there is probably another sensitive period in the development of Coping Intelligence, this is the period of early adulthood and middle age, when professional activity and family creation are associated with various difficulties, which can trigger prolonged inflammation in the body. This assumption requires additional research.
A possible neurochemical dynamic model of the Coping Intelligence is shown in fig. 3. A stressful situation leads to the activation of the sympathoadrenal system (SAS). The release of catecholamines (adrenaline, norepinephrine) provides a quick mobilization of the body and prepares it for action (“fight or flight”). Adrenaline initiates the release of insulin, which enhances glucose uptake by actively functioning organs. As a result of coping with stress, the concentration of adrenaline in the blood drops, as it is absorbed by the muscles (“utilized” adrenaline). However, if the body does not perform active actions and a high level of adrenaline remains in the blood, then the hypothalamic-pituitary-adrenal (HPA) system is triggered. The release of glucocorticoids (cortisol, etc.) maintains a high energy level in the body.
It should be noted that the studies do not take into account the “strength/weakness” of the nervous system and the severity of modal-specific activity. At the same time, the threshold of sensitivity in people with a “strong” and “weak” nervous system (highly active and low active in terms of V.M. Rusalov) differs (Ratanova, 2008). Under the influence of physical stress in respondents with a “strong” nervous system, the concentration of cortisol significantly decreases, while in respondents with a “weak” nervous system, it significantly increases. Respiratory rate normalization in respondents with a “strong” nervous system, compared with respondents with a “weak” nervous system, is achieved more slowly, they report the cessation of physical activity more quickly, but after additional stimulation they have more resources to continue it (Plachinta, 1978). The second (“cortisol”) period of the stress reaction is associated with a long-term mobilization of the body in response to the stressor, leading to suppression of the activity of the immune and reproductive systems, slowing down the development of the body. The suppression of regenerative processes continues during the sluggish stress response (Nikolaeva, 2019, p. 100). Termination of the stress response is ensured by the fact that activated glucocorticoid receptors reduce the activity of the HPA system with the help of negative biofeedback. Chronically low or high levels of glucocorticoids indicate suboptimal adaptation; a moderate or controlled increase in glucocorticoids is about physical and mental health (Nikolaeva, 2019, p. 103). The endogenous opioid regulatory system
(EOS) is aimed at producing a high level of endogenous opioids, thus inhibiting the development of a stress state (Bakhchina, 2022).
Acute stress state, in addition to the release of catecholamines, glucocorticoids (Vindaz et.al., 2018) and endogenous opioids, is associated with an increase in the level of Il-10, neutrotrophic factor (BDNF) (Linz, Puhlmann, Apostolakou et. al., 2019; Szuhany, Bugatti, Otto, 2015), high dopamine and serotonin levels (Aston-Jones, Chiang, Alexinsky, 1991; Kameneva et. al., 2022; Matsumoto & Hikosaka, 2009). Recovery of neurochemical balance after coping with acute stress occurs at low, medium, and high rates. The low rate of restoration of the neurochemical balance is associated with increased noadrenaline, a shift in the daily dynamics of glucocorticoid hormones (Bakhchina, 2022), decreased BDNF (Cattaneo et. al., 2016), increased Il-6 and TNF α (Dinan, 2009), and overexpression of ADR2A (Hohoff et. al., 2013), increased inflammation and a shift in the microbiota (Miller & Raison, 2016; Passos et. al., 2015; Straud, 2017), which determine the regressive line of human development in stressful situations. A moderate or controlled increase in glucocorticoids correlates (Nikolaeva, 2019) with an average rate of recovery of neurochemical parameters (stable line of development). The rapid restoration of neurochemical parameters ensures the progressive development of a person in stressful situations, due to the “reacted” adrenaline, restoration of cortisol to baseline, and a decrease in BDNF (Linz, Puhlmann, Apostolakou et. al., 2019). It should be noted the lack of empirical data on the state of the microbiota and the concentration of interleukins when a person quickly recovers from stressful situations. Based on the foregoing, we put forward a Neurochemical Dynamic Model of Coping Intelligence, which is presented in Fig. 3. The empirical verification of this model is the goal of our further research.
Highlights:
- The congruence of Stress Coping System levels ensures a high Coping Intelligence.
- The stress response is manifested in the HPA axis activation, elevation in pro-inflammatory cytokines and BDNF.
- Significant gaps in knowledge remain about neurochemistry of resilience.
- Resilience is associated with rhythms, cycles and synchronization of SCS.
- The effects of Early Life Stress can be offset by a healthy gut microbiota.
CRediT author statement: All authors contributed to the article and approved the submitted version.
Agorastos, A. & Chroustos, G.P. (2022). The neuroendocrinology of stress: the stress-related continuum of chronic disease development. Molecular Psychiatry, 27, 502-513. https://doi.org/10.1038/s41380-021-01224-9
Adjimann, T.S., Argañaraz, C.V. & Soiza-Reilly, M. (2021). Serotonin-Related Rodent Models of Early-Life Exposure Relevant for Neurodevelopmental Vulnerability to Psychiatric Disorders. Translational Psychiatry,11, 280. https://doi.org/10.1038/s41398-021-01388-6
Bahchina, A.V. (2022). Psychophysiology of stress. In: Psychophysiology: Ed. Yu.I. Aleksandrov, 5th edition. SPb.: Peter, 362-380. [Bahchina, A.V. (2022). Psihofiziologiya stressa. V: Psihofiziologiya: pod red. Aleksandrova Yu.I. 5-e izdanie. SPb.: Piter. 2022. 528 s.]
Brown, H. & Esterházy, D. (2021). Intestinal immune compartmentalization: implications of tissue specific determinants in health and disease. Mucosal Immunology,14, 1259–1270. https://doi.org/10.1038/s41385-021-00420-8
Cattaneo, A., Cattane, N., Begni, V. Pariante, C.M. & Riva M.A. (2016). The human BDNF gene: peripheral gene expression and protein levels as biomarkers for psychiatric disorders. Translational Psychiatry,6, 3-10. https://doi.org/10.1038/tp.2016.214
Danese, A. & Lewis, S.J. (2017). Psychoneuroimmunology of Early-Life Stress: The Hidden Wounds of Childhood Trauma? Neuropsychopharmacology,42, 99–114. https://doi.org/10.1038/npp.2016.198
Dinan, T.G. & Cryan, J.F. (2017). Microbes, Immunity, and Behavior: Psychoneuroimmunology Meets the Microbiome. Neuropsychopharmacology,42, 178–192. https://doi.org/10.1038/npp.2016.103
Egeland, M., Zunszain, P.A. & Pariante, C.M. (2015). Molecular mechanisms in the regulation of adult neurogenesis during stress. Nature Reviews Neuroscience,16, 189–200. https://doi.org/10.1038/nrn3855
Enck, P., Aziz, Q., Barbara, G., Farmer, A.D., Fukudo, S., Mayer, E.A., Niesler, B., Quigley, E.M.M., Rajilic-Stojanovic, M., Schemann, M., Schwille-Kiuntke, J., Simren, M., Zipfel, S. & Spiller, R.C. (2016). Irritable bowel syndrome. Nature Reviews Disease Primers,2, 16014. https://doi.org/10.1038/nrdp.2016.14
Faraji, J., Soltanpour, N., Lotfi, H., Moeeini, R., Moharreri, A-R., Roudaki, S., Hossenini, S.A., Olson, D.M., Abdollahi, A-A., Soltanpour, N., Mohajerani, M.H. & Metz, G.A.S. (2017). Lack of Social Support Raises Stress Vulnerability in Rats with a History of Ancestral Stress. Scientific Reports,7, 5277. https://doi.org/10.1038/s41598-017-05440-8
Feder, A., Nestler, E.J. & Charney, D.S. (2009). Psychobiology and molecular genetics of resilience. Nature Reviews Neuroscience, 10, 446-457. https://doi.org/10.1038/nrn2649
Gold, P. W. (2015). The organization of the stress system and its dysregulation in depressive illness. Molecular psychiatry, 20(1), 32-47.
Grayson, M. (2016). Irritable bowel syndrome. Nature, 533, 101. https://doi.org/10.1038/533S101a
Hill, M.N., Campolongo, P., Yehuda, R. & Patel, S. (2018). Integrating Endocannabinoid Signaling and Cannabinoids into the Biology and Treatment of Posttraumatic Stress Disorder. Neuropsychopharmacology, 43, 80–102 https://doi.org/10.1038/npp.2017.162
Hohoff, C., Gorji A., Kaiser, S., Willscher, E., Korsching, E., Ambree, O., Arolt, V., Lesch, K-P., Sachser. N., Deckert, J. & Lewejohann, L. (2013). Effect of Acute Stressor and Serotonin Transporter Genotype on Amygdala First Wave Transcriptome in Mice. PLoS ONE, 8(3), 1-13. https://doi.org/10.1371/journal.pone.0058880
Holz, N. E., Tost, H., & Meyer-Lindenberg, A. (2020). Resilience and the brain: a key role for regulatory circuits linked to social stress and support. Molecular psychiatry, 25(2), 379-396. https://doi.org/10.1038/s41380-019-0551-9
Kredlow, A.M., Fenster, R.J., Laurent, E.S., Ressler, K.J., & Phelps, E. (2022). Prefrontal Cortex, Amygdala, and Threat Processing: Implications for PTSD. Neuropsychopharmacoogy,47, 247–259. https://doi.org/10.1038/s41386-021-01155-7
Kuvaeva, I.O. & Volkova, E.V. (2022) The multidimensional nature of Coping Intelligence. In: History, modernity and prospects for the development of psychology in the system of the Russian Academy of Sciences: Proceedings of the International Anniversary Scientific Conference dedicated to the 50th anniversary of the Institute of Psychology of the Russian Academy of Sciences, November 16–18, 2022, Moscow / Ed. ed. D. V. Ushakov, A. L. Zhuravlev, A. V. Makhnach, N. E. Harlamenkova, A. V. Yurevich, I. I. Vetrova. – M.: Publishing house “Institute of Psychology of the Russian Academy of Sciences”. 500-502. [Kuvaeva, I.O. &Volkova, E.V. (2022). Mnogomernaya priroda sovladayushchego intellekta / Istoriya, sovremennost’ i perspektivy razvitiya psihologii v sisteme Rossijskoj Akademii nauk: Materialy Mezhdunarodnoj yubilejnoj nauchnoj konferencii, posvyashchennoj 50-letiyu sozdaniya Instituta psihologii RAN, 16–18 noyabrya 2022 g., Moskva / Otv. red. D. V. Ushakov, A. L. ZHuravlev, A. V. Mahnach, N. E. Harlamenkova, A. V. YUrevich, I. I. Vetrova. – M.: Izd-vo «Institut psihologii RAN», 500- 502.]
Laird, K.T., Krause, B., Funes, C. & Lavretsky H. (2019). Psychobiological factors of resilience and depression in late life. Translational Psychiatry,9, 88. https://doi.org/10.1038/s41398-019-0424-7
Linz, R., Puhlmann, L.M.C., Apostolakou, F., Mantzou, E., Papassotiriou, I., Chrousus, C.P., Engert, V. & Simger, T. (2019). Acute psychosocial stress increases serum BDNF levels: an antagonistic relation to cortisol but no group differences after mental training. Neuropsychopharmacology,44, 1797–1804 https://doi.org/10.1038/s41386-019-0391-y
Lopez, M., Ruiz, M.O., Rovnaghi, C.R., Tam, G.K-Y., Hiscoux, J., Gotlib, I.M., Barr, D.A., Carrion, V.G. & Anand, K.J.S. (2021). The social ecology of childhood and early life adversity. Pediatric Research, 89, 353–367. https://doi.org/10.1038/s41390-020-01264-x
Malhi, G.S., Das, P., Bell, E., Mattingly, G. & Mannie, Z. (2019). Modelling resilience in adolescence and adversity: a novel framework to inform research and practice. Translational Psychiatry, 9, 316. https://doi.org/10.1038/s41398-019-0651-y
McEwen, B.S., Bowles, N.P., Gray, J.D., Hill, M.N., Hunter, R.G., Karatsoreos, I.N. & Nasca, C. (2015). Mechanisms of stress in the brain. Nature Neuroscience, 18, 1353-63. https://doi.org/10.1038/nn.4086
McEwen, B., Nasca, C. & Gray, J. (2016). Stress Effects on Neuronal Structure: Hippocampus, Amygdala, and Prefrontal Cortex. Neuropsychopharmacology,41, 3–23. https://doi.org/10.1038/npp.2015.171
Ménard, C., Pfau, M.L., Hodes, G.E. & Russo, S.J. (2017). Immune and Neuroendocrine Mechanisms of Stress Vulnerability and Resilience. Neuropsychopharmacology,42, 62–80. https://doi.org/10.1038/npp.2016.90
Miller, A.H. & Raison, C.L. (2016). The role of inflammation in depression: from evolutionary imperative to modern treatment target. Nature Reviewers Immunology, 16, 22-34. doi: 10.1038/nri.2015.5.
Nikolaeva, E.I. (2019). Psychophysiology: Textbook for universities. third generation standard. SPb.: Peter. 704 p. [Nikolaeva E. (2019). Psihofiziologiya: Uchebnik dlya vuzov. Standart tret’ego pokoleniya. SPb.: Piter. 704 s.]
Passos, I.C., Vasconcelos-Moreno, M.P., Costa, L.G., Kunz, M., Brietzke, E., Quevedo, J., Salum, G., Magalhaes., P.V., Kapczinski, F. & Kauer-Sant’Anna M. (2015). Inflammatory markers in post-traumatic stress disorder: a systematic review, meta-analysis, and meta-regression. Lancet Psychiatry, 2, 1002–1012. doi: 1016/S2215-0366(15)00309-0
Pittenger, C. & Duman, R. (2008). Stress, Depression, and Neuroplasticity: A Convergence of Mechanisms. Neuropsychopharmacology, 33, 88–109. https://doi.org/10.1038/sj.npp.1301574
Plachinta, M. A. (1978). The strength of the nervous system and the nature of autonomic reactions in untrained people during physical activity of different intensity. Questions of Psychology, 4, 37-42. [Plachinta, M. A. (1978). Sila nervnoj sistemy i harakter vegetativnyh reakcij u netrenirovannyh lyudej pri fizicheskoj nagruzke raznoj intensivnosti. Voprosy psihologii, 4, 37-42.]
Price, R.B. & Duman, R. (2020). Neuroplasticity in cognitive and psychological mechanisms of depression: an integrative model. Molecular Psychiatry, 25(3), 530-543. doi: 10.1038/s41380-019-0615-x.
Raison, C. & Miller, A. (2017). Pathogen–Host Defense in the Evolution of Depression: Insights into Epidemiology, Genetics, Bioregional Differences and Female Preponderance. Neuropsychopharmacology, 42, 5–27. https://doi.org/10.1038/npp.2016.194
Rakesh, G., Morey, R.A., Zannas, A.S., Malik, Z., Marx, C.E., Clausen, A.N., Kritzer, M.D. & Szabo, ST. (2019). Resilience as a translational endpoint in the treatment of PTSD. Molecular Psychiatry, 24, 1268-1283. doi: 10.1038/s41380-019-0383-7.
Ratanova, T.A. (2008). Psychophysical scaling. Power of sensations, power of the nervous system and sensitivity. Moscow. Second edition. [Ratanova, T.A. (2008). Psihofizicheskoe shkalirovanie. sila oshchushchenij, sila nervnoj sistemy i chuvstvitel’nost’. Moscow. 2 izdanie].
Schwartz, M. & Shechter, R. (2010). Protective autoimmunity functions by intracranial immunosurveillance to support the mind: The missing link between health and disease. Molecular Psychiatry,15, 342–354. https://doi.org/10.1038/mp.2010.31
Sterling, P. (2004). Principles of allostasis: optimal design, predictive regulation, pathophysiology, and rational therapeutics. In: Allostasis, gomeostasis, and the costs of psychological adaptation. Ed. By J. Schulkin, 17 – 64.
Sterling, P. & Eyer, J. (1988). Allostasis: A New Paradigm to Explain Arousal Pathology. In: Handbook of life stress, cognition and health / Edited by S.Fisher and J. Reason. John Wiley & Sons, 629-649.
Straub, R. H. (2017). The brain and immune system prompt energy shortage in chronic inflammation and ageing. Nature Reviewers Rheumatology,13, 743–751. https://doi.org/10.1038/nrrheum.2017.172
Vandael, D. & Gounko, N.V. (2019). Corticotropin releasing factor-binding protein (CRF-BP) as a potential new therapeutic target in Alzheimer’s disease and stress disorders. Transational Psychiatry, 9, 272. https://doi.org/10.1038/s41398-019-0581-8
Vindas, M.A., Fokos, S., Pavlidis, M., Höglund, E., Dionysopoulou, S., Ebbesson, L.O.E., Papandroulakis, N., & Dermon, C.R. (2018). Early life stress induces long-term changes in limbic areas of a teleost fish: the role of catecholamine systems in stress coping. Scientific Reports,8, 5638. https://doi.org/10.1038/s41598-018-23950-x
Wittig, R.M., Crockford, C., Weltring, A., Langergraber, K.E., Deschner, T. & Zuberbuhler, K. (2016). Social support reduces stress hormone levels in wild chimpanzees across stressful events and everyday affiliations. Nature Communications, 7, Article number 13361, 1-8. https://doi.org/10.1038/ncomms13361
Zannas, A.S., Wiechmann, T., Gassen, N.C. & Binder, E.B. (2016). Gene–Stress–Epigenetic Regulation of FKBP5: Clinical and Translational Implications. Neuropsychopharmacology,41, 261–274. https://doi.org/10.1038/npp.2015.235